 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Occurrence of "Nuages" and "Lamellae Anulata" during Spermatogenesis in Dermatobia hominis (Diptera: Cuterebridae) Irani Quagio-Grassiotto, Edy de Lello Departamento de Morfologia, Instituto de BiociEncias, UNESP, 18618-000 Botucatu, SP, Brasil
Code Number: OC95108
Size of Files:
Text: 25K
Graphics: Photos (jpg) 548K / Halftones (gif) 595K
Various types of "nuages" and "lamellae anulata" can be found
during Dermatobia hominis spermatogenesis. In
spermatogonia, the "nuages" occur as granules juxtaposed to
the cytoplasmic face of the nuclear envelope or as cytoplasmic
granules similar to glycogen granules. In spermatocytes, in
addition to the "nuages", dense spherical bodies of
approximately 1.0 um in diameter are also observed. In the
spermatids the "nuages" can be of the following types:
perinuclear granules, spherical granules with diameters
varying in length from 0.5 to 1.0 um, granules similar to
glycogen granules, granules with variable diameters which
accumulate at the flagellum base forming the centriole
adjunct, or remain in the cytoplasm. "Nuages" can also be
observed in these cellular types as dense masses, without a
definite outline and are common to animal germinal cells in
general. The "lamellae anulata" on the other hand, are
observed only in spermatocytes I and in early spermatids,
being always immersed in electron-dense material of indefinite
outline. In spermatids, the "lamellae anulata" are close to
the nuclear envelope suggesting, in spite of opposing
opinions, that these cells are envolved in the synthesis and
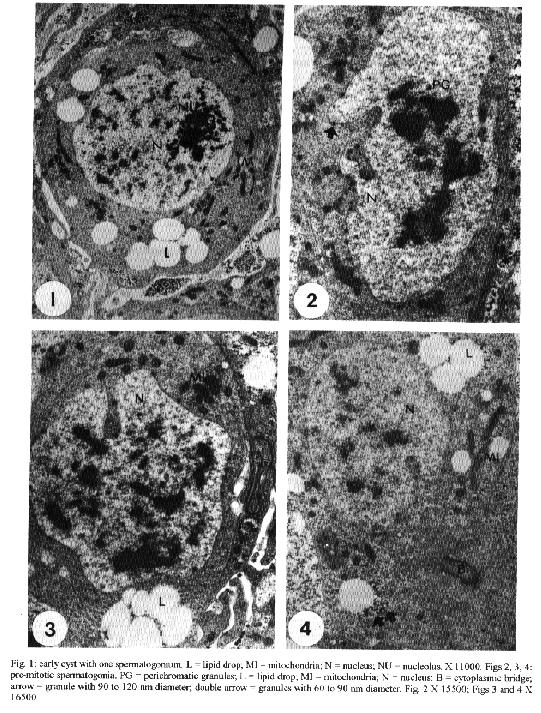
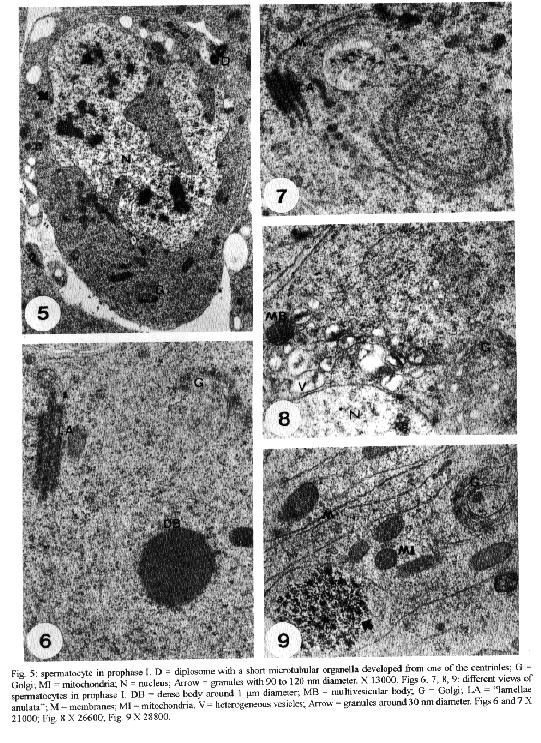
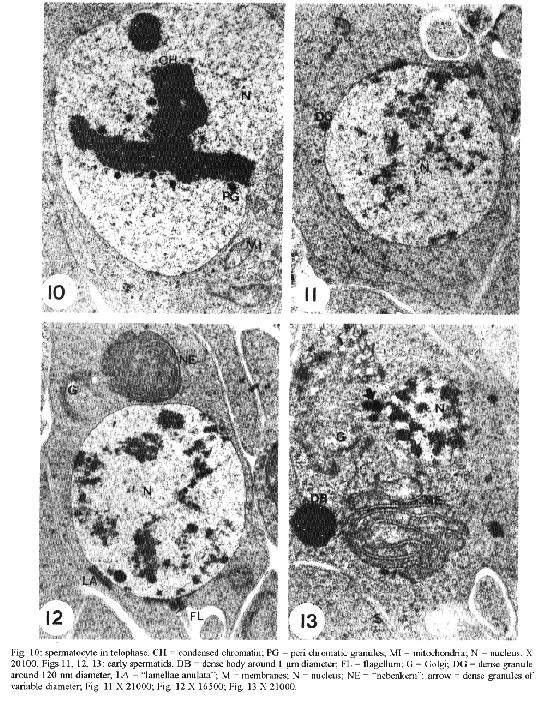
transport of material from the nucleus to the cytoplasm.Key words: Dermatobia hominis - spermatogenesis - "nuage" - "lamellae anulata" Cytoplasmic electron-dense materials not surrounded by membranes are a characteristic of animal germinal cells, generically known as "nuages". They occur in different sizes and shapes, receive different names and are present during the whole life cycle of these cells. They are abundant during oogenesis and spermatogenesis, contain RNA, appear isolated in the cytoplasm close to the nuclear envelope and are associated with mitochondria (Eddy 1975) or with "lamellae anulata" (Kessel 1983). "Lamellae anulata" are transient organelles, composed of stacks of cisterns having pores, similar to nuclear pores. They are found in several cells, especially in animal germinal cells, being well developed in oocytes (Kessel 1983, 1992). Despite the innumerable efforts their function so far is unknown (Merisko 1989, Kessel 1992). We studied the different types of "nuages" observed during the spermatogenesis of Dermatobia hominis, their morphology, their relationship with cellular organelles and their period of occurrence, and we compared them to the ones described in the male germinal cells of other animals. Our aim is to contribute to a better understanding of them. MATERIALS AND METHODS For this study, pupae were developed in the laboratory from larvae supplied by highly infected bovines. The animals used in this experiment were confined overnight in stables with concrete floors covered by a wooden grid, with 7 cm spacing. Mature larvae abandoned the hosts and fell to the ground during the early morning hours; they were protected from being stamped upon by the spaces in the wooden grid. Each morning larvae were collected, taken to the laboratory and separated by weight; those weighing between 500 to 600 mg developed male imagos (Lello 1979). They were then placed in plastic boxes containing a 6 to 8 cm layer of humid soil where they penetrated to pupate. Boxes were kept at 25 C, with 70-80% relative humidity in a BOD incubator (Fanem, Brasil) during the entire pupal stage which lasted from 30 to 34 days. From the very beginning of the pupal stage, pupae were removed daily from the soil and dissected in a saline solution for the removal of the insects' gonads; these were preserved in 2.5% glutaraldehyde solution with phosphate buffer, at a 0.1 M, pH of 7.3, for 2.5 hr, and then placed in 1% osmium tetroxide with the same buffer, for 2 hr, in a darkened area, and embedded in araldite. The ultrathin sections were highlighted using a saturated solution of uranyl acetate in 50% alcohol and with lead citrate (Reynolds 1963); observations and photo micrographs were made using a Phillips EM 301 transmission electron microscope. RESULTS At the time of cyst formation the definitive spermatogonia (Hannah-Alava terminology 1965) in D. hominis, mitochondria and lipid droplets were present in the cytoplasm that were poor in other organelles; nuclei were circular, nucleoli had a well-developed granulated region, almost reaching the nuclear envelope (Fig. 1). Various perichromatic granules from 90 to 120 nm in diameter were observed in the nuclei during interphasic periods. Similar granules were also observed in the cytoplasm, next to a deep penetration of the nuclear envelope (Fig. 2). Mitochondria were either isolated or organized in clusters having an electron-dense material in the center (Fig. 3), or associated with lipid droplets (Fig. 4). Other strongly electron-dense granules with a 60 to 90 nm diameter, were randomly distributed in the cytoplasm or associated with lipid droplets (Fig. 4). Like spermatogonia, spermatocytes at the beginning of the meiotic prophase I, had granules varying from 90 to 120 nm in diameter close to the cytoplasmic face of the nuclear envelope. Short microtubular organelles developed from the centrioles (Fig. 5). Besides this there was an extensive system of membranes to which the "lamellae anulata" were associated (Fig. 7). The "lamellae anulata" were formed by at most five cisterns, with pores similar to nuclear pores. They were linear, organizing themselves in stacks, and appearing immersed in an electron-dense material without a definite outline (Figs 6, 7). In addition to the "lamellae anulata" other spherically dense bodies, about 1.0 um in diameter and clusters of electron-dense particles, about 30 nm in diameter, appeared in the cytoplasm (Fig. 9). A well-developed Golgi apparatus, randomly dispersed mitochondria, multivesicular bodies and vesicles with a heterogeneous content (Figs 6, 8, 9) were also observed. The only type of "nuage" observed during the entire meiosis, was a cluster of particles about 30 nm in diameter. By the end of telophase II, the nuclei of the newly formed cells assumed a spherical shape, and their still condensed chromatin appeared surrounded by perichromatic granules varying in length from 90 to 120 nm (Fig. 10). At the beginning of their differentiation the spermatids presented spherical nuclei and diffused chromatin. Electron-dense granules, of about 120 nm in diameter, could be occasionally observed opposed to the cytoplasmic face of the nuclear envelope (Fig. 11). Like the spermatocytes, the spermatids presented "lamellae anulata". They were linear, continuous with the endomembrane system, being formed at most by three stacks of cisterns close to the nuclear envelope; they were immersed in an electron-dense material with an indefinite outline, reaching from the nuclear pores to the lamellar pores (Fig. 12). The presence of a well-developed Golgi apparatus, a cluster of mitochondria composing the "nebenkern", early flagellum, elongated membranes, and randomly dispersed spherically dense bodies with diameters varying in lenght from 0.5 to 1.0 um and others varying in length from 100 to 300 nm were also characteristic of these cells (Figs 11, 12, 13). During the initial period of spermatid elongation the cytoplasm contained clusters of particles, about 40 nm in diameter, a deposit of electron-dense material around the base of the flagellum (the centriole adjunct) and occasional electron-dense granules, approximately 0.2 by 0.4 um in length attached to the nuclear envelope (Figs 14, 15). The material around the flagellum base increased, forming a dense mass intermingled with irregular spaces filled with cytoplasm (Figs 16, 17). Multivesicular bodies and vesicles of heterogeneous content were found in the cytoplasm (Fig. 19). Multivesicular bodies were also at the caudal end of the elongated spermatids (Fig. 18). DISCUSSION The electron-dense granules, varying from 90 to 120 nm in diameter observed in the spermatogonia, spermatocytes I and initial spermatids of D. hominis associated to the pore region of the nuclear envelope, had dimensions and electron density similar to the perichromatic granules found in the nuclei. The position occupied by such granules and their similarity to the perichromatic granules suggested the occurence of an intense process of synthesis and transport of material from the nucleus to the cytoplasm. Russel and Frank (1978) described rat spermatogonia and spermatocytes granules of the same dimension and electron density, located next to the Golgi or between this and the nuclear envelope. Electron-dense granules associated to the nuclear envelope of the spermatocytes I were also observed by Fuge (1976) in Pales ferruginea (Diptera: Tipulidae) and by Kessel in Drosophila melanogaster (Diptera: Drosophilidae); the granules were composed of RNA and proteins (Fuge 1976, Kessel 1981) and therefore, this is in accordance with the supposition that they must be the nuclear material being transferred to the cytoplasm. The electron-dense material, observed among clusters of mitochondria in the cytoplasm of D. hominis spermatogonia, is a common feature in animal germinal cells (Eddy 1975). However, its occurrence during spermatogenesis in insects was only reported for D. melanogaster spermatocytes (Kessel 1981). This material, known as intermitochondrial cement, presents various types of RNAs, proteins, cytochromes and lipids (Toury et al. 1977). The fact that most of these proteins present the same electrophoretic mobility in polyacrylamide gel as mitochondrial proteins, led Toury et al. (1977) to the conclusion that their presence was associated to the origin, development and biogenesis of these organelles. The present knowledge of the interaction between the mitochondrial and nuclear genoma and the processes of synthesis and transference of the mitochondrial proteins to its interior (Darnell et al. 1990, Alberts et al. 1994), brought a new and strong support to their supositions. Cruz-Landim (1979) studying Myogryllys sp. (Orthoptera) spermiogenesis observed that mitochondria appeared associated to the nuclear envelope, before the development of "nebenkern". This fact also led her to suggest the transference of material from the nucleus, which would participate in mitochondrial differentiation during spermiogenesis. Another kind of electron-dense granule observed during D. hominis spermatogenesis was approximately 30 nm in diameter and occured in the form of clusters in the cytoplasm of the spermatocytes and early spermatids. Similar granules were previously reported by Russel and Frank (1978) in rat spermatocytes; according to these authors they were not ribosomes because unlike to the ribosomes, they reacted intensely to ferrocyanide. Russel and Frank (1978) also eliminated the possibility that these granules be glycogen granules because they differed in morphology from the glycogen granules described by Russel and Burguet (1977) in rat Leydig cells. Our experience in D. hominis led us to believe that they really were glycogen granules. We share this supposition with Wolf et al. (1987), who described similar granules in the spermatocytes of Orgya thyellina. However, proper cytochemical tests for the validation of this supposition must be done. The electron-dense spherical bodies, approximately 1.0 um diameter, observed in the spermatocytes I and in the early spermatids of D. hominis are not similar to the "nuages" described by Russel and Frank (1978). However, such bodies could correspond to the "spongious bodies" observed by them in the spermatocytes I of rats, or to the "chromatoid body satellite" described by Fawcett et al. (1970) in hamster spermatocytes, or to the "dense cytoplasmic masses" observed by Kessel (1981) in D. melanogaster early spermatids, or even to the "lamellar bodies" reported by Henning and Kremer (1990) in D. hydei spermatocytes and early spermatids. They certainly correspond to the electron-dense material, described in early spermatids of Melipona quadrifasciata anthidioidis, as "chromatoid body" by Cruz-Landin et al. (1981). In our opinion, the electron-dense spherical bodies would be composed of RNA and proteins, just like the dense cytoplasmic masses (Kessel 1981) and the "lamellar bodies" (Henning & Kremer 1990). The electron-dense material that accumulates around the base of the flagellum in D. hominis early spermatids and in most of the early spermatids of insects is known as "centriole adjunct" (Gatenby & Tahmisian 1959, Phillips 1970, Baccetti 1972, Yasuzumi 1974). The development of centriole adjunct in Acrida lata spermatids described by Yasuzumi et al. (1970) suggests that the dense bodies that at first appear in the cytoplasmic face of the nuclear envelope contribute to the centriole formation. According to Cruz-Landim (1979) these dense bodies were the result of an aggregation of ribosomes. The aspect and position of the centriole adjunct in the early spermatids in insects are similar to those of the chromatoid body in early mammaliam spermatids (Sud 1961, Eddy 1970, 1974, Fawcett et al. 1970, Comings & Okada 1972, Yasuzumi, 1974, Russel & Frank, 1978). The chromatoid body in mammals is an electron-dense structure, with an irregular outline, a fibro-granular aspect, in the cytoplasm, appearing during the meiotic prophase and which, in the early spermatids, migrates to the base of the flagellum. Chemical analysis of the centriole adjunct showed that it contains RNA and protein (Yasuzumi et al. 1970, Taffarel & Esponda 1980), detected previously in chromatoid bodies in mammals (Sud 1961, Eddy 1970, Soderstrom & Parvinen 1976). For this reason, we believe that the centriole adjunct of insects and the chromatoid bodies in mammals are equivalent cellular structures, as already suspected, but refuted by Yasuzumi et al. (1970). We also observed in D. hominis spermatocytes during prophase I and in early spermatids, another kind of "nuage", that occured as a cloud of electron-dense material associated with "lamellae anulata". In his various studies about "lamellae anulata", Kessel (1981, 1985) reported the same type of association in D. melanogaster. Kessel (1981) observed that in early spermatids "lamellae anulata" differentiate in the interior of what he described as "dense cytoplasmic masses". He also demonstrated that these dense masses correspond to the perinuclear material found in spermatocytes I, which, in the spermatids, change their position and relocate in the cytoplasm. Furthermore, Kessel (1981) demonstrated that the masses are made up of granular and fibrillar subunits comparable to the nucleolar subunits, to the material associated to the nuclear pores and to the poliribosomes, possibly being formed by RNA and proteins. Kessel (1981) based on the rhythm of the RNA and protein synthesis during spermatogenesis in Drosophila sp. (Olivieri & Olivieri 1965, Henning 1967, Gould-Somero & Holland 1974), and his own observations, suggested that the dense cytoplasmic masses of early spermatids contain genic products of long duration such as rRNAs and mRNAs. He also proposed the hypothesis that the activation of these products in the spermatids should be associated with the appearance of "lamellae anulata" or specifically to the pores of these lamellae. In a further study, Kessel (1985), demonstrated that in D. melanogaster spermatocytes I, the "lamellae anulata" which had a perinuclear location differentiate in the interior of a fibrogranular material, of possible nucleolar origin, aligned continuosly from the nuclear to the lamellar pores. In D. hominis, the electron-dense material associated with the "lamellae anulata" did not have a definite outline and was observed in the cytoplasm of spermatocytes I, without a fixed localization. In early spermatids it was arranged in a continuous manner between the nuclear envelope and the "lamellae anulata". This arrangement led us to assume that there was a recent synthesis of this material, perhaps by the spermatid. This contradicts previous data regarding the post meiotic synthesis of RNA during spermatogenesis in insects (Olivieri & Olivieri 1965, Henning 1967, Brink 1968, Gould-Somero & Holland 1974). However, it finds support in data about other animal species (Monesi 1965, Moore 1975, Kierszenbaum & Tres 1975, Iatrou et al. 1978, Monesi et al. 1978, Hecht et al. 1986, Oliva et al. 1988, Klemm et al. 1989). Our proposition is corroborated by Muckenthaler (1964) in Melanoplus differentialis and more recently by Huyser et al. (1990) in D. hydei. ACKNOWLEDGEMENTS To EM Laboratory of IB, Botucatu, for the facilities; to Mr Antonio Vicente Salvador for technical help and Mrs Sandra Aparecida Andrades da Silva for the typewritting. REFERENCES Alberts B, Bray D, Lewis J, Raff M, Roberts K, Watson JD 1994. Molecular biology of the cell, 3rd ed. New York and London, Garland Publishing, Inc. 1294 pp. Baccetti B 1972. Insect sperm cells. Adv Insect Physiol 9: 315-397. Brink NC 1968. Protein synthesis during spermatogenesis in Drosophila melanogaster. Mutat Res 5: 192-194. Comings DE, Okada TA 1972. The chromatoid body in mouse spermatogenesis: evidence that it may be formed by the extrusion of nucleolar components. J Ultrastruct Res 39: 15-23. Cruz-Landim C 1979. Ultrastructural characterization of nuage in spermatocytes of Myogryllus sp. (Orthoptera). The nucleus 22: 67-74. Cruz-Landim C, Moraes RIM, Beig D 1981. Caracterizacao do material electron denso de origem nuclear, durante a espermiogEnese de abelhas (Hymenoptera, Apidae). Rev Bras Biol 4: 421-426. Darnell J, Lodish H, Baltimore P 1990. Molecular cell Biology. 2nd ed. New York, Scientific American Books, W.H. Freeman and Company, 1005 pp. Eddy EM 1970. Cytochemical observations on the chromatoid body of the male germ cells. Biol Reprod 2: 114-128. Eddy EM 1974. Fine structural observations on the form and distribution of nuage in germ cell of the rat. Anat Rec 178: 731-757. Eddy EM 1975. Germ plasm and the differentiation of the germ cell line. Int Rev Cytol 43: 229-280. Fawcett DW, Eddy EM, Phillips DM 1970. Observation on the fine structure and relationships of the chromatoid body in mammalian spermatogenesis. Biol Reprod 2: 129-153. Fuge H 1976. Ultrastructure of cytoplasmic nucleolus-like bodies and nuclear RNP particles in late prophase of tipulid spermatocytes. Chromosoma 56: 363-379. Gatenby JB, Tahmisian TN 1959. Centriole adjunct, centrioles, mitochondria and ergastoplasm in orthopteran spermatogenesis. An electron microscopy study. Cellule 60: 104-134. Gould-Somero M, Holland L 1974. The timing of RNA synthesis for spermiogenesis in organ cultures of Drosophila melanogaster testes. Wilhelm Roux Arch Entwicklungsmech 174: 133-148. Hannah-Alava A 1965. The premeiotic stages of spermatogenesis. Adv Genet 13: 157-226. Hecht NB, Bower PA, Waters SH, Yelick PC, Distel RJ 1986. Evidence for haploid expression of mouse testicular genes. Exp Cell Res 164: 183-190. Henning W 1967. Untersuchungen zu struktur und funktion des lampenbursten-y-chromosomes in der spermatogenese von Drosophila. Chromosoma 22: 294-357. Henning W, Kremer H 1990. Spermatogenesis of Drosophila hydei. Int Rev Cytol 123: 129-175. Huyser P, Beckers L, Top B, Hermans M, Sinke R, Henning W 1990. Poly [d (C - A)] poly [d (G - T)] is highly transcribed in testes of Drosophila hydei. Chromosoma 100: 48-55. Iatrou K, Spira AW, Dixon GH 1978. Protamine messenger RNA: evidence for early synthesis and accumulation during spermatogenesis in rainbow trout. Dev Biol 64: 82-98. Kessel RG 1981. Origin, differentiation, distribution and possible functional role of annulate lamellae during spermatogenesis in Drosophila melanogaster. J Ultrastruct Res 75: 72-96. Kessel RG 1983. The structure and function of annulate lamellae: porous cytoplasmic and intranuclear membrane. Int Rev Cytol 82: 181-303. Kessel RG 1985. The relationships of annulate lamellae, fibrogranular bodies, nucleolus and polyribosomes during spermatogenesis in Drosophila melanogaster. J Ultrastruct Res 91: 183-191. Kessel RG 1992. Annulate lamellae: a last frontier in cellular organelles. Int Rev Cytol 133: 43-120. Kierszenbaum AL, Tres L 1975. Structural and transcriptional features of the mouse spermatid genome. J Cell Biol 65: 258-270. Klemm U, Lee C, Burfeind P, Hake S, Engel W 1989. Nucleotide sequence of a DNA encoding rat protamine and the haploid expression of the gene during rat spermatogenesis. Biol Chem Hoppe-Seyer 370: 293-301. Lello E 1979. Desenvolvimento pos embrionario das gonadas de Dermatobia hominis (Linnaeus Jr., 1781) (Diptera: Cuterebridae). Thesis, IBBMA-UNESP, 73 pp. Merisko EM 1989. Annulate lamellae: an organelle in search of a function. Tissue & Cell 21: 343-354. Monesi V 1965. Synthetic activities during spermatogenesis in the mouse. Exp Cell Res 39: 197-224. Monesi V, Geremia R, D'agostino A, Boitani C 1978. Biochemistry of male germ cell differentiation in mammals: RNA synthesis in meiotic and postmeiotic cells. Curr Top Dev Biol 12: 11-37. Moore GPM 1975. DNA-dependent RNA synthesis in fixed cells during spermatogenesis in mouse. Exp Cell Res 68: 462-465. Muckenthaler FA 1964. Autoradiographic study of nucleic acid synthesis during spermatogenesis in the grasshopper, Melanoplus differentialis. Exp Cell Res 35: 531-547. Oliva R, Mesquita J, Mesquita C, Dixon GH 1988. Haploid expression of the rooster protamine mRNA in the postmeiotic stages of spermatogenesis. Dev Biol 125: 332-340. Olivieri G, Olivieri A 1965. Autoradiographic study of nucleic acid synthesis during spermatogenesis in Drosophila melanogaster. Mut Res 2: 366-380. Phillips DM 1970. Insect sperm: their structure and morphogenesis. J Cell Biol 44: 243-277. Reynolds ES 1963. The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J Cell Biol 17: 208-212. Russel LD, Burguet S 1977. Ultrastructure of Leydig cells as revealed by secondary tissue treatment with a ferrocyanide osmium mixture. Tissue & Cell 9: 751-766. Russel L, Frank B 1978. Ultrastructural characterization of nuage in spermatocytes of the rat testis. Anat Rec 190: 79-98. Soderstrom K-O, Parvinen M 1976. Incorporation of [3H] uridine by the chromotoid body during rat spermatogenesis. J Cell Biol 70: 239-246. Sud BN 1961. The "chromatoid body" in spermatogenesis. Q J Micros Sci 102: 273-292. Taffarel M, Esponda P 1980. Fine-structural cytochemistry of the centriolar adjunct in grasshopper spermatids. Mikroskopie 36: 35-42. Toury R, Clerot JC, Andre J 1977. Les groupements mitochondriaux des cellules germinales des poissons teleosteens Cyprinides. IV. Analyse biochimique des constituants du "ciment" intermitochondrial isole. Biol Cell 30: 225-232. Wolf KW, Baumgart K, Traut W 1987. Cytology of Lepidoptera. II. Fine structure of eupyrene and apyrene primary spermatocytes in Orgya thyellina. Eur J Cell Biol 44: 57-67. Yasuzumi G 1974. Electron microscope studies on spermiogenesis in various animal species. Int Rev Cytol 37: 53-120. Yasuzumi G, Sugioka T, Tsubo I, Yasuzumi F, Matano Y 1970. Spermatogenesis in animals as related by electron microscopy. XX Relationship between chromatoid bodies and centriole adjunct in spermatids of grasshopper Acrida lata. Z Zellforsch Mikrosk Anat 110: 231-242.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95108d.gif] [oc95108c.gif] [oc95108a.gif] [oc95108b.gif]Photo images[oc95108b.jpg] [oc95108a.jpg] [oc95108d.jpg] [oc95108c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}