 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Identification of Anopheles (Nyssorhynchus) rondoni (Diptera: Culicidae) in Mato Grosso State, Brazil: An Analysis of Key Character Variability Richard C Wilkerson/+, Lawrence J Hribar*, Eric G Milstrey**, Guillermo Calderon Falero*** Department of Entomology, Walter Reed Army Institute of Research Washington, DC 20307, USA *Florida Medical Entomology Laboratory, 200 9th St. SE, Vero Beach, FL 32962 **485th Medical Detachment, Fort Polk, LA 71459 ***Pan American Health Organization, Brasilia, DF, Brasil
A morphological study was made of a population of Anopheles (Nyssorhynchus) rondoni (Neiva and Pinto) from northern Mato Grosso, Brazil. This population usually lacked the primary key character of a dark basal band on hindtarsomere 3, i.e., hindtarsomere 3 was all white as in most other members of the subgenus. It was determined that this species can be recognized instead by the presence of a dark spot on the thorax made up of a large dark prescutellar space that is contiguous with a concolorous central area on the scutellum. A secondary character of a dark area on the costa created by the fusion of the humeral dark, presector dark and sector dark proximal spots is also usually reliable. Regression analyses comparing the lengths and ratios of the dark bands on hindtarsomeres 2 to those on 3 describe a straight line relationship. This suggests that the 'atypical' population is at one end of a character gradient. We propose that in the subgenus Nyssorhynchus individuals that have a long basal band on hindtarsomere 2 are more likely to also have a basal band on hindtarsomere 3. The pupal stage of this species has not been previously described. Reared-associated specimens from this study show that the pupa can be easily differentiated from all other Nyssorhynchus by the relatively stout, usually 2 or 3 branched (1-5), setae 1 and 5 on segments IV-VII. Key words: Anopheles (Nyssorhynchus) rondoni - key characters - identification - pupa In spite of two recent revisions (Faran 1980, Linthicum 1988), identification of females of Anopheles (Nyssorhynchus) species is often difficult. This is because of an incomplete understanding of inter- and intraspecific variability, because of unresolved species complexes and because of lack of study specimens from all parts of species ranges. To identify the highly variable taxa belonging to this subgenus, it is therefore appealing to rely on presumably unambiguous “key” characters. One such character is the presence of a basal dark band on hindtarsomere 3 of An. (Nys.) rondoni (Neiva and Pinto). With the exception of An. (Nys.) nigritarsis Chagas and other naturally occurring variants (discussed below), this tarsomere is usually entirely white in subgenus Nyssorhynchus. We found that this “key” character was uncommon in a population of An. (Nys.) rondoni from Peixoto de Azevedo, Mato Grosso State, Brazil, and that this would likely lead to misidentification of these specimens as An. (Nys.) benarrochi Gabaldon, Cova Garcia and Lopez. We offer here an analysis of this and other characters used to distinguish this species and suggest an alternative means for identification. Numerical analysis showed a positive correlation between the extent of basal dark scales on hindtarsomere 2 and those on hindtarsomere 3. This allows us to predict that the occurrence of a dark band on hindtarsomere 3 is more likely in species that have a longer band on hindtarsomere 2. Also, this study provided the first examples of the pupa of this species for future morphological analysis. MATERIALS AND METHODS Female Anopheles were collected from human bait at Peixoto de Azevedo, Mato Grosso, Brazil on 20 April, 1993 by two of us (E.G.M. and G.C.F.). These females were allowed to feed and then transported to the US Army Research Unit in Rio de Janeiro (USAMRU-B). They were maintained in a humid environment and traumatically induced to lay eggs by removal of a wing after 48-72 hr. Larvae were reared at approximately 25 C. Two or three adults of each sex with associated larval and pupal exuviae (Peixoto F1 females and males) from each of 26 progeny broods and their female parents (Peixoto P1 females) were retained for morphological study. A similar number of larvae and adults of both sexes were frozen for molecular studies. Specimens are deposited in the National Museum of Natural History (NMNH), Smithsonian Institution, Washington, DC.; U.S. Army Medical Research Unit, Rio de Janeiro; Instituto Oswaldo Cruz, Rio de Janeiro; and the Nucleo de Pesquisa Taxonomica e Sistemática em Entomologia Medica, at the Universidade de Sao Paulo. The Peixoto de Azevedo specimens bear the reference code BR Rio 020 followed by the progeny brood number in parentheses, e.g. BR Rio 020 (19). The following progeny broods are included in this study: (19-21), (25-45), (48) and (53). Other specimens studied are older holdings of the National Museum of Natural History, Smithsonian Institution (NMNH specimens). They are from Corrientes and Jujuy Provinces, Argentina and, Sao Paulo State and Costa Marques, Rondonia State, Brazil. Except for specimens from the latter locality they were used by Faran (1980) in developing his descriptions and keys. Measurements were made using an SMZ-10 microscope with a camera lucida and a Summagraphics SummaSketch Model MM1201 using 'INPAD' software written by Joseph Russo (Office of Information Management, Smithsonian Institution). Wing spot names have been converted from the names used by Faran (1980) to the system proposed by Wilkerson and Peyton (1990), abbreviated as follow (Fig. 1): prehumeral dark (PHD); humeral pale (HP); humeral dark (HD); presector pale (PSP); presector dark (PSD); sector pale (SP); sector dark (SD) [the sector dark is divided into two spots, the sector dark proximal (SDP) and sector dark distal (SDD) when an accessory sector pale (ASP) spot is present]; subcostal pale (SCP); and preapical dark (PD). Equivalent names in Faran (1980) are: basal dark = PHD; humeral pale = HP; subbasal dark = HD; subbasal pale = PSP; presectoral dark = PSD; presectoral pale = SP; sectoral dark = SDP; sectoral pale = ASP; subcostal dark = SDD; and preapical dark = PD. Vein nomenclature follows Harbach and Knight (1980). Veins M1+2, M1, and M2 correspond to veins M, M1+2 and M3+4.in Faran (1980) We observed the presence/absence of the above wing spots and measured their lengths. We also measured the lengths of hindtarsomeres 2 and 3 and lengths of the basal bands on hindtarsomeres 2 and 3. We then calculated ratios of HP to PHD, SCP to SDD, and proportion of hindtarsomeres 2 and 3 dark. All ratios were transformed by X' = (X + 3/8)1/2 (Kihlberg et al. 1972) prior to analysis. Possible differences among NMNH specimens, Peixoto P1 females and F1 males and F1 females were assessed by analysis of variance (SAS GLM procedure), and mean separations were performed by the Ryan-Einot-Gabriel-Welsch multiple range test. Differences in frequency of missing wing spots PSP, SP, ASP and dark or light scaling of wing vein M1 were analyzed by applying the likelihood ratio chi-square (SAS FREQ procedure) to 2 X 4 contingency tables. This test was chosen due to small cell sizes for some of the variables. The number of observations for a given group (e.g., Peixoto P1 females) was not equal for all characters due to damage to specimens, rubbing of wing spots, etc. All statistical analyses were conducted with the aid of the Statistical Analysis System (SAS Institute 1985). Ratios of basal dark bands on hindtarsomeres 2 and 3 were calculated and plotted. Several morphological characters were used to verify that the 26 progeny broods from Mato Grosso State, Brazil, were An. (Nys.) rondoni. The male genitalia of an individual from each brood was examined and found to agree with Faran's (1980) genitalia characters, i.e., the ventral lobes with setae usually on the basal lobule not extending to the base of the apicolateral lobe and the preapical plate very weakly sclerotized and ill-defined. The male genitalia of An. (Nys.) strodei Root is quite similar but has the setae of the ventral lobes on the lateral margins usually reaching the base of the apicolateral lobe and the preapical plate is weakly to moderately sclerotized and moderately well-defined. Anopheles (Nys.) rondoni is different from strodei in two other characters. First, the pale wing scales are white to occasionally white mixed with very pale yellow in An. (Nys.) strodei but pale yellow or pale yellow with white on the costal veins in An. (Nys.) rondoni. Second, rondoni exhibits a distinctive, contrasting, dark prescutellar space, a character not known to us in strodei, or any other species of the subgenus.
RESULTS This study evaluates characters used by Faran (1980) to identify An. (Nys.) rondoni. His key characters were: (1) hindtarsomere 3 dark in basal 0.20-0.35; (2) vein C predominantly dark (Fig. 1B), HD, PSD and SDP spots fused into one large spot (i.e. PSP and SP spots missing); and (3) PD of vein M1+2 extending uninterrupted onto vein M1. We evaluated these and other characters to test the likelihood of misidentification of An.(Nys.) rondoni which lack a band on hindtarsomere 3.
TABLE 1. Basal dark band on the hindtarsomeres 2 and 3, Anopheles (Nys.) rondoni National museum of National History specimens (NMNH) and male and female progeny of wild females from Peixoto de Azevedo, Mato Grosso, Brazil.
------------------------------------------------------------
Range/Mean STD Legs/ % with
individuals bands
------------------------------------------------------------
Hindtarsomere 2
NMNH females 0.64-0.89 0.77 0.07 30/27 100%
Peixoto P1 females 0.47-0.69 0.57 0.05 24/24 100%
Peixoto F1 females 0.42-0.77 0.59 0.66 97/49 100%
Peixoto F1 males 0.39-0.74 0.59 0.07 92/47 100%
Hindtarsomere 3
NMNH females 0.19-0.42 0.29 0.05 30/26 100%
Peixoto P1 females 0.0-0.19 0.11a 0.05a 24/24 19%
Peixoto F1 females 0.0-0.22 0.12a 0.06a 96/49 29%
Peixoto F1 males 0.0-0.27 0.14a 0.07a 92/47 27%
-------------------------------------------------------------
a of those with a bandTABLE II Morphological characters of museum specimens, wild-caught mothers, and female progeny of Anopheles (Nys.) rondoni. Means +/- 1 standard error within rows followed by the same letter are not different (Ryan- Einot-Gabfiel-Welsch multiple range test)
-------------------------------------------------------------
Peixoto F1 Peixoto F1 Peixoto P1
females males females
-------------------------------------------------------------
1HT 3 dark (mm) 0.021+/-0.005b 0.023+/-0.004b 0.015+/-0.007b
HT 2 dark (mm) 0.511+/-0.006b 0.498+/-0.006b 0.522+/-0.013b
2Ratio HT3 dark 0.116+/-0.009b 0.140+/-0.011b 0.112+/-0.026b
Ratio HTII dark 0.594+/-0.009b 0.594+/-0.007b 0.569+/-0.011b
HT 3 length (mm)0.603+/-0.005c 0.600+/-0.004c 0.640+/-0.013b
HT 2 length (mm)0.861+/-0.006c 0.838+/-0.005c 0.919+/-0.014b
PHD (mm) 0.039+/-0.003b 0.034+/-0.003b 0.065+/-0.005a
SCP (mm) 0.187+/-0.005a 0.151+/-0.003b 0.173+/-0.009a,b
DSD (mm) 0.642+/-0.006b,c 0.625+/-0.006c 0.679+/-0.009b
--------------------------------------------------------------
NMNH F value df P
females
--------------------------------------------------------------
1HT 3 dark (mm) 0.216+/-0.010a 198.25 3,238 0.0001
HT 2 dark (nun) 0.811+/-0.026a 145.03 3,239 0.0001
2Ratio HT3 dark 0.291+/-0.010a 63.22 3,85 0.0001
Ratio HTII dark 0.768+/-0.018a 62.85 3,239 0.0001
HT 3 length (mm) 0.737+/-0.013a 53.28 3,238 0.0001
HT 2 length (mm) 1.050+/-0.019a 93.65 3,239 0.0001
PHD (mm) 0.036+/-0.006b 3.31 3,215 0.021
SCP (mm) 0.145+/-0.014b 8.53 3,238 0.0001
DSD (mm) 0.754+/-0.035a 15.20 3,202 0.0001
------------------------------------------------------------
1: includes those without a band on hindtarsomere 32: includes only those with a band on hindtarsomere 3 TABLE III. Frequency tabulation of presence or absence of wing spots; presector pale (PSP), sector pale (SP), accessory sector pale (ASP) and preapical dark spot of vein M1+2 with or without pale scales at base of M1
------------------------------------------------------------
PSP SP
Present Absent Present Absent
------------------------------------------------------------
NMNH 6(20%) 24(80%) 0 30(100%
females
Peixoto P1 0 20(100%) 1(5%) 19(95%)
females
Peixoto F1 13(13.3%) 85(86.7%) 1(1.1%) 97(98.9)
females
Peixoto F1 4(4.3%) 90(95.7%) 0 94(100%)
males
------------------------------------------------------------
ASP M1
Present Absent Pale scaled All dark
------------------------------------------------------------
NMNH 8(26.7%) 22(73.3%) 8(27.6%) 21(72.4%)
females
Peixoto P1 21(100%) 0 5(41.7%) 7(58.3%)
females
Peixoto F1 92(93.9%) 6(6.1%) 45(47.9%) 49(52.1%)
females
Peixoto F1 91(96.8%) 3(3.2%) 54(62.1%) 33(37.9%)
females
------------------------------------------------------------
Vein C (Table III) - Faran (1980) described vein C as predominantly dark because of the fusion of the subbasal, presectoral and sectoral dark spots (= HD, PSD and SDP spots), i.e., the PSP and SP spots were missing.
-----------------------------------------------------------
PSP SP ASP NMNH females Peixoto F1 Peixoto P1 Peixoto FI
females mothers males
-----------------------------------------------------------
0 0 0 19(63.3%)[3] 6(6.1%) 0 3(3.2%)[1]
0 0 1 5(17.7%) 78(79.6%)[7] 19(95%) 87(92.6%)[5]
0 1 1 0 1(1) 1(5%)[1] 0
1 0 0 3(10%)[1] 0 0 0
1 0 1 3(10%) 13(13.3%) 0 4(4.3%)
0 1 0 0 0 0 0
1 1 1 0 0 0 0
1 1 0 0 0 0 0
Wings 30 98 20 94
Individuals26 49 20 47
------------------------------------------------------------
0=absent; 1=present; number (percent) [no. also lacking
subcostal pale spot]; PSP=presector pale spot; SP=sector pale
spot; ASP=accessory sector pale spot.
There were significantly different percentages of Peixoto F1 females, F1 males, P1 females, and NMNH specimens lacking the PSP (X2 = 12.178, df = 3, P < 0.014) and ASP (X2 = 79.018, df = 3, P < 0.0001) wing spots. No differences in frequencies of missing wing spots among female progeny, male progeny, mothers, and museum specimens were found for the SP spot (X2 = 5.329, df = 3, P < 0.149). Of the eight possible combinations of these three pale spots (Table IV) the majority of wings from Peixoto de Azevedo (85.7% of F1 females, 95.8% of F1 males and 95% of P1 females) exhibited an absence of the PSP and SP spots (classes 000 and 001); most specimens, but not all, therefore agreeing with Faran (1980). Among the NMNH specimens, 81% agreed with Faran (1980) (63.3% were missing all three spots, 17.7% had the ASP), the remainder did not agree. Few Peixoto specimens (6.1% of females and 2.1% of males) had all three pale spots missing. Some individuals, in addition to missing most or all of the above pale spots, also lacked the subcostal pale (SCP), thus giving the costa of the wing an even darker appearance. Overall, the NMNH specimens were therefore darker on this part of the wing than the Peixoto population. The occurrence of pale spots appears to be non-random since several classes of spots were rare or did not occur. For instance, PSP and SP absent, ASP present (001) is the most common class for Peixoto specimens but PSP absent, SP present, ASP absent (010) was not represented, even though both classes have only one of the spots missing. The ratio of the HP to the PHD spots and the ratio of the SCP to the SDD spots (Table V) were also calculated because of their usefulness in distinguishing other Oswaldoi Subgroup species. The PHD spot was missing or relatively short in comparison to the HP in An. (Nys.) rondoni, which is consistent with Faran (1980); however, the ranges of both the NMNH specimens and Peixoto specimens for the ratio HP/PHD, 1.95 - 10.48 and 1.04 - 14.2 respectively, exceed that given by Faran (1980) (2.0-3.3). The SCP is relatively short or missing, also consistent with FaranÆs (1980) figure of rondoni. With only one exception, the SCP is less than 0.5 the size of the DSD. A specimen with a SCP that is >0.5 of the DSD could be interpreted as being An. (Nys.) rangeli Gabaldon, Cova Garcia and Lopez. No statistically significant differences between groups were found for the following variables: ratio of HP and PHD spots (F = 1.08; df = 3, 154; P > 0.3557); ratio of SCP and SDD wing spots (F = 4.55; df = 3, 202; P > 0.05); and length of HP wing spot (F = 0.70; df = 3, 154; P > 0.5563). TABLE V. Ratios of the prehumeral dark spot to the humeral pale spot (PHD/HP) and subcostal pale spot to the distal sector dark spot (SCP/DSD) in Anopheles (Nys.) rondoni from specimens in the National Museum of Natural History, Smithsonian Institution (NMNH) and from Peixoto de Azevedo, Mato Grosso, Brazil (020)
--------------------------------------------------------
Range Mean STD No. wings
--------------------------------------------------------
HP/PHD
NMNH females 1.95-10.48 3.94 1.87 19
Peixoto P1 females 1.78-5.76 3.06 1.16 8
Peixoto F1 females 1.42-10.24 4.11 1.79 70
Peixoto F1 males 1.04-14.2 4.35 2.32 61
SCP/SDD
NMHM females 0.16-0.33 0.25 0.06 8
Peixoto P1 females 0.14-0.35 0.26 0.06 21
Peixoto F1 females 0.09-0.52 0.30 0.07 91
Peixoto F1 males 0.09-0.47 0.26 0.08 86
--------------------------------------------------------
Preapical dark spot (PD) of vein M - Faran (1980) noted that
An. (Nys.) rondoni differed from other species in the
Oswaldoi Group by having no pale spot on M1+2 and M1 where
M1+2 branches into M1 and M2. We found this to be partially
true (Table III). If this character is strictly interpreted
then there should not be pale scales on M1 where it branches
from M1+2. We found pale scales on the posterior portion of
M1 in a large proportion of our sample. However, we never
noted pale scales on the anterior portion of this vein. A
survey of other Nyssorhynchus in the collection of the NMNH
showed that the pale spot on both the anterior and posterior
portions of the vein usually to be present; but specimens
similar to rondoni with dark scales on the anterior part of
the wing were found in An. (Nys.) strodei of the
Albimanus Section, and An. (Nys.) marajoara Galvao and
Damasceno and An. (Nys.) braziliensis (Chagas) of the
Argyritarsis Section. A significant frequency of a darker M1
(all dark vs dark anterior with pale scales on posterior
portion of the vein) was found in the museum specimens
compared to Peixoto mothers and progeny (X^2 = 11.383, df = 3,
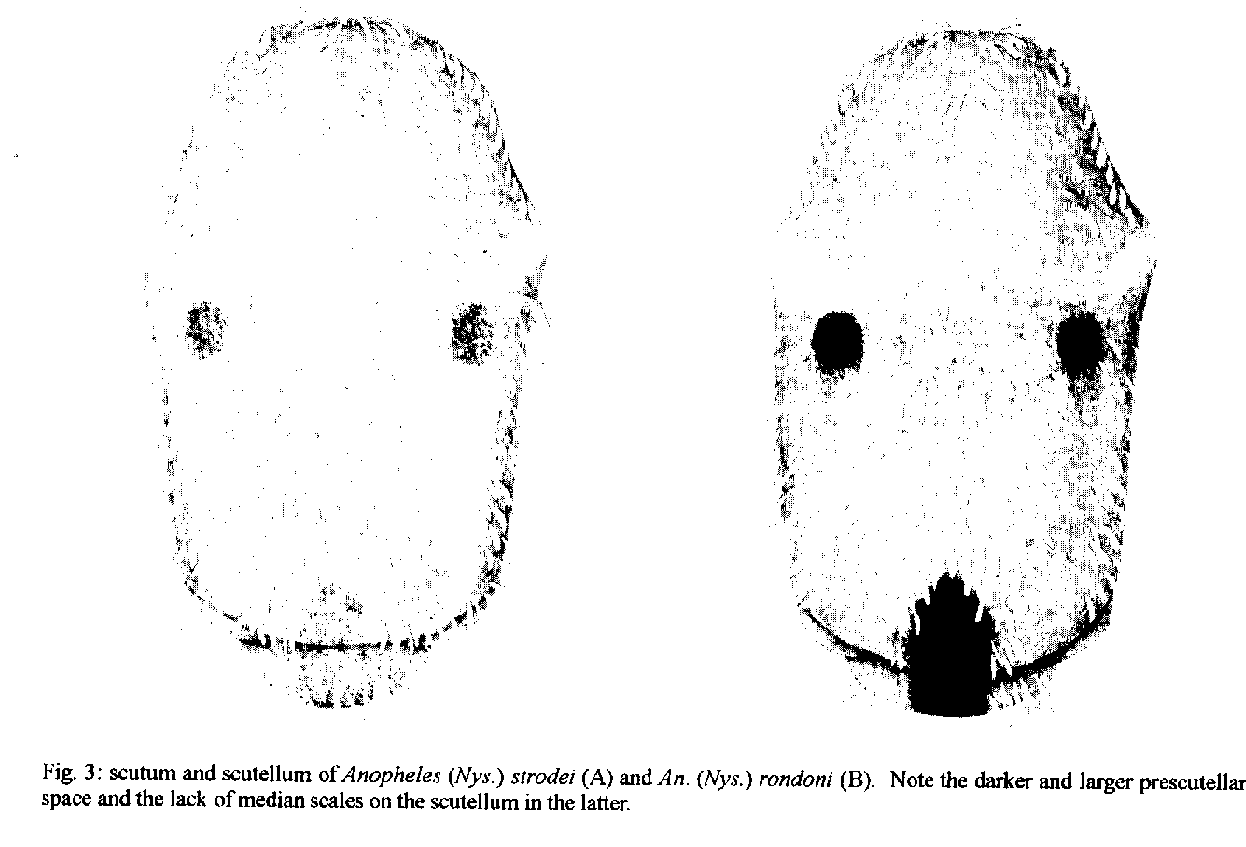
P < 0.01).Prescutellar space - In FaranÆs (1980) discussion and in the original description (Neiva & Pinto 1922) it is noted that the presence of a very dark brown to black, subtriangular, prescutellar space distinguishes this species. This dark area is contiguous with a concolorous central area on the scutellum, and together form a conspicuous spot (Fig. 3). We found this character to be present in all An. (Nys.) rondoni examined and absent in other Nyssorhynchus. In addition, the central dark area on the scutellum lacks scales, a condition not known to us in other Nyssorhynchus.
Fig. 3: scutum and scutellum of Anopheles (Nys) strodei (A) and An. (Nys.) rondi (B). Note the darker and larger prescutellar space and the lack of median scales on the scutellum in the later.
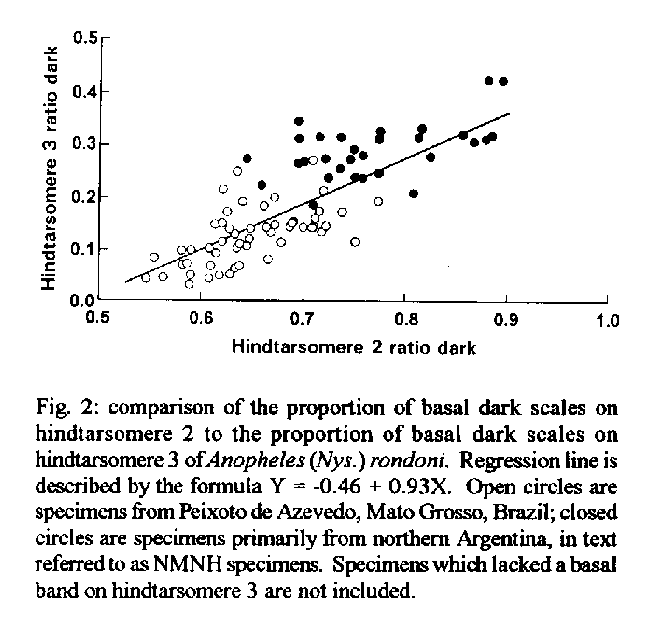
DISCUSSION The discovery of a population of An. (Nys.) rondoni not exhibiting the primary “key” character of a dark basal band on hindtarsomere 3 requires a reevaluation of characters used to recognize this species. If a specimen of rondoni has a basal band on hindtarsomere 3 it can be easily identified, unless one of the variants of other Albimanus Section species, that also have this band, are encountered at the same time (see below). The two other characters used by Faran (1980) in his key to separate An. (Nys.) rondoni from other members of the Oswaldoi Group are likewise not constant. In his key one could be led to an identification of An. (Nys.) aquasalis Curry or An. (Nys.) benarrochi with the following combination of characters: hindtarsomere 3 pale, PSP or SP present, M1 with pale scales, base of hindtarsomere 2 0.3-0.6 dark, and SCP less than 0.5 length of the SDD. Anopheles (Nys.) aquasalis can be eliminated on the basis of various characters found in the description, and because it is not known to be sympatric with An. (Nys.) rondoni. Anopheles (Nys.) benarrochi, though, is similar in many ways and occurs with rondoni. We suggest that the best character for recognition of adults of rondoni therefore is the large, dark, prescutellar space and contiguous dark area, lacking scales, on the scutellum (Fig. 3). The secondary key character of a dark area on vein C resulting from the fusion of the HD, PSD and SDP was also usually in agreement with Faran (1980). The case of An. (Nys.) rondoni individuals lacking a basal dark band on hindtarsomere 3 is similar to instances of variability in other Nyssorhynchus species which, instead, gain bands on hindtarsomere 3 and sometimes 4. Varieties, species or variants have been recognized in the following: An. (Nys.) nigritarsis (a valid species with diagnostic characters of basal bands on hindtarsomeres 3 and 4); An. (Nys.) albimanus Wiedemann (vars. bisignatus Hoffman and trisignatus Hoffmann) [Hoffmann 1938, Rozeboom 1963, Faran 1980]; An. (Nys.) aquasalis vars. guarauno Anduze and delta Anduze and the synonym deltaorinoquensis Cova Garcia, Pulido F. and Amanista M.) [Anduze 1948, Faran 1980]; An. (Nys.) strodei (variant) [Rachou and Ferraz 1951]; An. (Nys.) triannulatus (Neiva and Pinto) (synonym Cellia cuyabensis [Neiva and Pinto]) [Pinto 1938, Galvao and Lane 1941]; An. (Nys.) darlingi Root (variant) [Harbach et al. 1993]; and An. (Nys.) albitarsis (synonym imperfectus Correa and Ramos) [Correa and Ramos 1943, Galvao 1943]. In all of the above, the band on hindtarsomere 2 is relatively long compared to other Nyssorhynchus species. They are as follow (after Faran 1980 and Linthicum 1988): An. (Nys.) strodei, 0.30 - 0.50; An. (Nys.) darlingi, 0.35 - 0.55; An. (Nys.) albitarsis, 0.60 - 0.90; An. (Nys.) albimanus, 0.40 - 0.80; An. (Nys.) triannulatus, 0.40 - 0.70; and An. (Nys.) aquasalis, 0.30 - 0.60. One might predict that the extra band on hindtarsomere 3 will be found in other species with a relatively long hindtarsomere 2 band, e.g. An. (Nys.) galvaoi Causey, Deane and Deane, An. (Nys.) benarrochi, An. (Nys.) pictipennis (Philippi) or An. (Nys.) deaneorum Rosa-Freitas. Regression analysis of the ratios of dark on hindtarsomere 2 vs hindtaromere 3, and regression analysis of the actual measurements, supports the hypothesis that it is more likely that there will be a dark basal band on hindtarsomere 3 if a larger proportion of hindtarsomere 2 is dark (Fig. 2). The statistical comparisons between Peixoto de Azevedo and NMNH specimens show some significant differences, i.e., incidence of PSP and ASP spots, dark scales on M1, and the lengths and proportions of the basal dark on hindtarsomeres 2 and 3. The differences in these morphological characters could be evidence of separate species, but they probably simply represent seasonal or geographic variation. Any attempt to establish separate species status for the Peixoto specimens must await further collections, since the Peixoto and NMNH collections are almost at opposite ends of the known distribution of the species. The differences in tarsomere banding may be an expression of some pressure that is either seasonal or geographic (Le Sueur & Sharp 1991, Kitthawee et al. 1992, Le Sueur et al. 1992) which contribute to darkening on the wing as well. If keys for the identification of Nyssorhynchus species rely only on leg characters, it would be possible to assign “nontypical” specimens to the wrong subgenus, i.e. Argyritarsis Section species with basal bands on hindtarsomeres 2-5 could be mistaken for subgenus Kerteszia. We do not believe this to be a problem, however, since numerous other characters exist to separate Nyssorhynchus from Kerteszia (Peyton et al. 1992). Linthicum (1988) uses the acrotstichal and dorsocentral areas having numerous scales to separate Nyssorhynchus, and Wilkerson and Strickman (1990) use scutal markings and wing spots. This report of a common, easily misidentified variant of what formerly was considered to be a readily identifiable species, exemplifies problems of identification of species in the subgenus Nyssorhynchus. Relatively little is known about inter- and intraspecific variation in Nyssorhynchus, or how to identify the numerous cryptic species. Without an accurate understanding of these taxa, results of epidemiological, ecological and control studies can be easily confused. This is a clear illustration of the need for further extensive collection and analysis of the species in the subgenus Nyssorhynchus, whose member species are responsible for the majority of malaria parasite transmission in the Neotropics. Acknowledgments To Jose Bento Lima, for laboratory assistance in Brazil. To Taina R Litwak for the preparation of Figs 1 and 3, Bonnie K Pattok for preparing Fig. 2 and Jayson I Glick, E L Peyton and Terry A Klein for their helpful reviews of the manuscript. REFERENCES Anduze PJ 1948. Notas entomologicas I. Dos variedades nuevas de A. aquasalis. Bol Med (Caracas) 1: 17-19. Correa RR, Ramos AS 1943. Descricao de uma nova sub-especie de anofelino do sub-genero Nyssorhynchus Blanchard, 1902 (Diptera, Culicidae). Rev Paulista Med 22: 246-248. Faran ME 1980. Mosquito studies (Diptera: Culicidae). XXXIV. A revision of the Albimanus Section of the subgenus Nyssorhynchus of Anopheles. Contrib Am Entomol Inst (Ann Arbor) 15: 1-215. Galvao ALA 1943. Chaves para a determinacao das especies do subgenero Nyssorhynchus do Brasil. Arq Hig Saude Publica 8: 141-162. Galvao ALA, Lane J 1941. Observacoes sobre alguns anofelinos de Salobra, Mato Grosso (Diptera, Culicidae). Rev Biol Hyg 11: 10-18. Harbach RE, Knight KL 1980. TaxonomistsÆ glossary of mosquito anatomy. Plexus, Marlton, N.J. Harbach RE, Roberts DR, Manguin S 1993. Variation in the hindtarsal markings of Anopheles darlingi (Diptera: Culicidae) in Belize. Mosq Syst 25: 192-197. Hoffmann CC 1938. La formacion de razas en los Anopheles mexicanos. II. Anopheles albimanus y sus variedades en la Republica Mexicana. An Inst Biol Univ Nac Auton Mex 9: 167-180. Kihlbeg JK, Herson JH, Schultz WE 1972. Square root transformation revisited. J Roy Statist Soc Ser C Appl Statist 21: 76-81. Kitthawee S, Edman JD, Upatham ES 1992. Anopheles dirus size and fecundity: relationship to larval density and protein accumulation. Southeast Asian J Trop Med Public Health 23: 128-131. Le Sueur D, Sharp BL 1991. Temperature-dependent variation in Anopheles merus larval head capsule width and adult wing length: implications for adult taxonomy. Med Vet Entomol 5: 55-62. Le Sueur D, Sharp BL, Appleton CC 1992. Dark-scaled areas on adult Anopheles mosquitoes are selectively affected by temperature-related size variation. Med Vet Entomol 6: 396-398. Linthicum KJ 1988. A revision of the Argyritarsis Section of the subgenus Nyssorhynchus of Anopheles (Diptera: Culicidae). Mosq Syst 20: 99-271. Neiva A, Pinto C 1922. Contribuicao para o conheci-mento das anophelinas do estado de Matto Grosso com a descripcao de uma nova especie. Braz - Med 36: 321-322. Peyton EL, Wilkerson RC, Harbach RE 1992. Comparative analysis of the subgenera Kerteszia and Nyssorhynchus of Anopheles (Diptera: Culicidae). Mosq Syst 24: 51-69. Pinto C 1938. Disseminacao da malaria pela aviacao; biologia de Anopheles gambiae e outros anofelinos do Brasil. Mem Inst Oswaldo Cruz 34: 293-430. Rachou RG, Ferraz DM 1951. Contribuicao ao conhecimento da distribuicao geografica dos anofelinos no Brasil: estado de Santa Catarina. Rev Bras Malariol Doencas Trop 3: 540-554. Rozeboom LE 1963. Mutant forms of Anopheles albimanus Wiedemann (Diptera: Culicidae). Proc Entomol Soc Wash 65: 110-114. SAS Institute. Inc 1985. SAS user's guide: statistics. Ver. 5th edition. Cary, NC. Wilkerson RC, Strickman D 1990. Illustrated key to the female anopheline mosquitoes of Central America and Mexico. J Am Mosq Control Assoc 6: 7-34. Wilkerson RC, Peyton EL 1990. Standardized nomenclature for the costal wing spots of the genus Anopheles and other spotted-wing mosquitoes (Diptera: Culicidae). J Med Entomol 27: 207-224. Partial support for this work was provided by NIH grant AI-31034 to L. Philip Lounibos, Florida Medical Entomology Laboratory, University of Florida, Vero Beach. The views of the authors do not purport to reflect the views of the supporting agencies. +Corresponding author. Present address: Museum Support Center, Smithsonian Institution, Washington, DC 20560 Received 5 January 1995 Accepted 12 May 1995
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95116a.gif]Photo images[oc95116a.jpg]Line drawing images[oc95116c.gif] [oc95116b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}