 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem. Inst. Oswaldo Cruz, Rio de Janeiro, Vol. 90 (5), sep./oct. 1995 Ultrastructural Aspects of Spermatogenesis in Metamicrocotyla macracantha (Alexander,1954) Koratha,1955 (Monogenea; Microcotylidae)
Maria de Fatima Diniz Baptista Farias, Anna Kohn/+, Ortrud Monika Barth*/+ Laboratorio de Helmintos Parasitos de Peixes, Departamento de Helmintologia *Laboratorio de Ultraestrutura Viral, Departamento de Virologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
Code Number: OC95120
Size of Files:
Text: 15
Graphics: Photos (jpg) 606K / Halftones (gif) 721K
Ultrastructural aspects of spermatogenesis, spermiogenesis and of the mature
spermatozoon of a microcotylid monogenean Metamicrocotyla macracantha
parasite from Mugil liza, are described. The irregularly-shaped
spermatogonia divides by successive mitoses, forming the primary
spermatocytes, identified by the presence of synaptonemal complexes in their
nuclei. The spermatids formed by meiotic cell divisions of the secondary
spermatocytes, differentiate into a mature spermatozoon. Cross sections of
the head and the middle region of mature spermatozoa show the nucleus with
strong condensed chromatin, the mitochondria with short cristae, peripheral
microtubules and two axonemes with a 9+1 pattern, confirming the
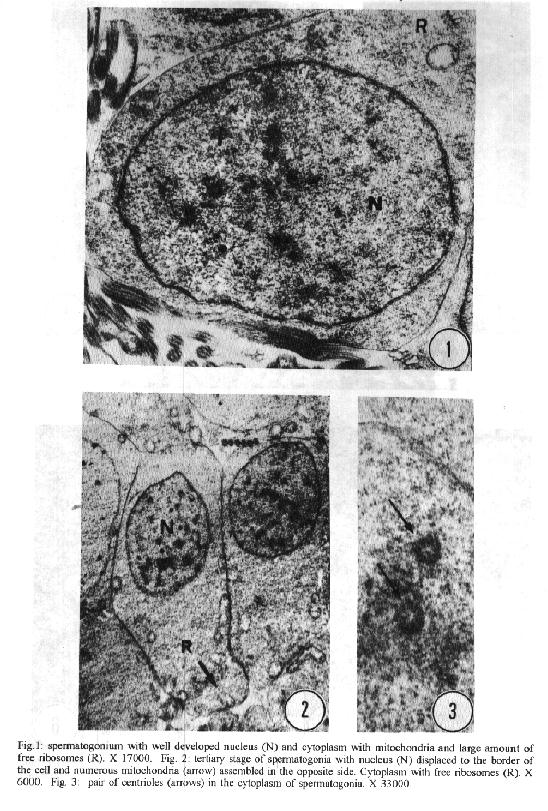
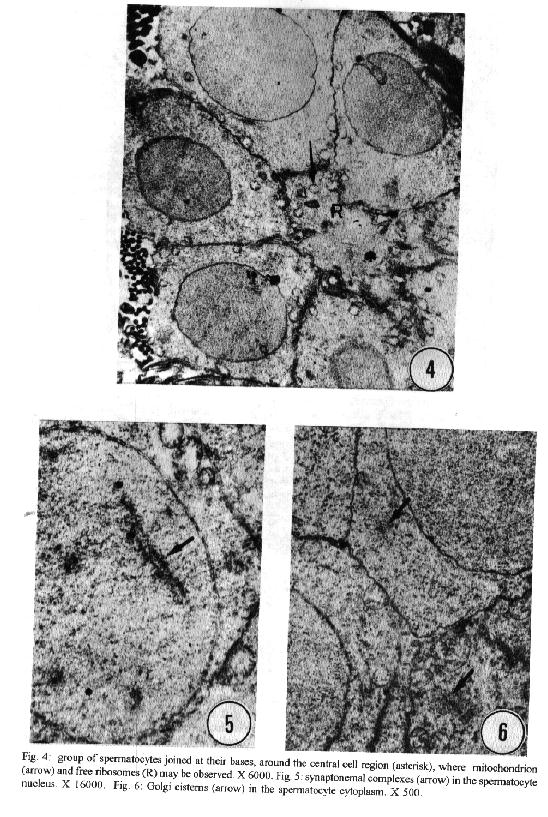
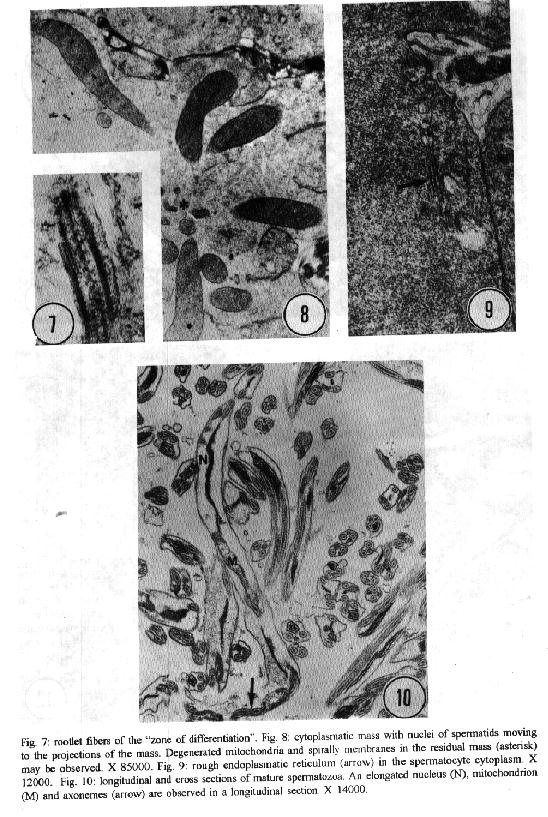
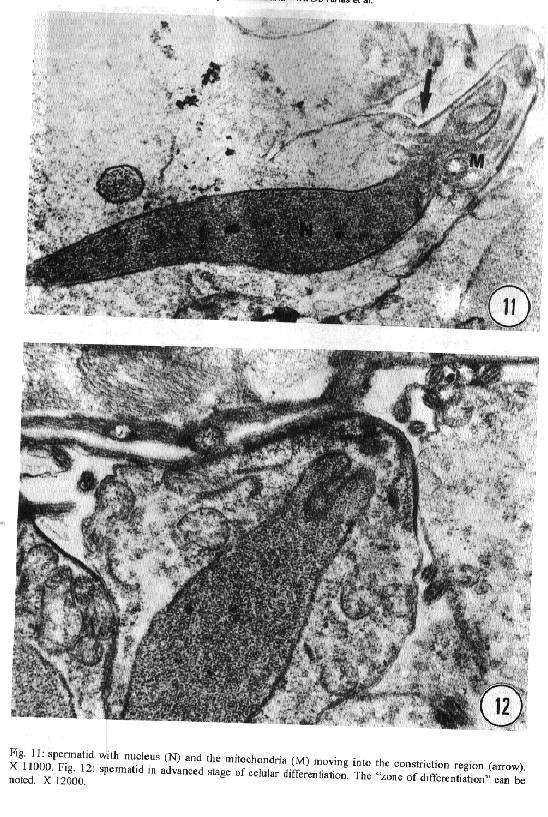
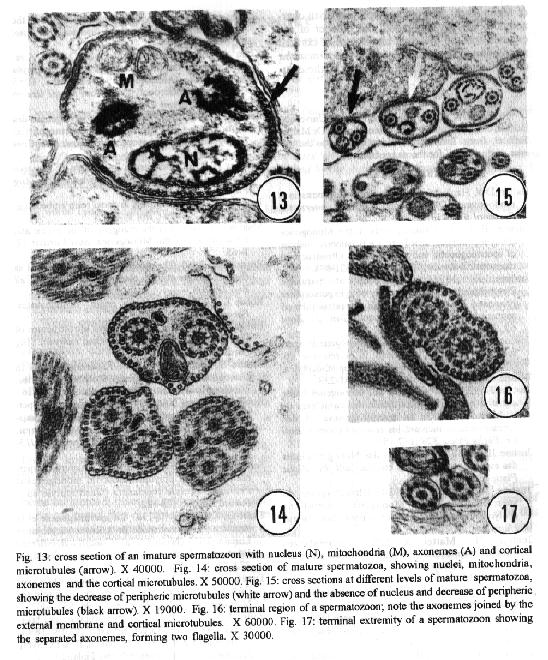
characteristics of this genus. Key words: ultrastructure - Metamicrocotyla macracantha - Monogenea - spermatogenesis - spermiogenesis - spermatozoa The additional data obtained on spermiogenesis and the ultrastructural aspects of spermatozoa are complementary to the traditional methods of morphological analysis, even to molecular biology techniques. Associated with other characteristics, these data have been utilized for phylogenetic analysis of the monogenean by various authors (Lambert 1980, Justine et al.1985b, 1991a,b,1993a, Lebedev 1988, Malmberg 1990). In the present study, spermatogenesis and ultrastructural aspects of the spermatozoa of the oligonchoinean, Metamicrocotyla macracantha, are examined for the first time by transmission electron microscopy. MATERIALS AND METHODS Mugil liza (Mugilidae) were collected by fishermen at the 'Canal de Marapendi', coast of Rio de Janeiro, Atlantic Ocean. Infected gills were removed and examined in a saline medium under stereoscopic microscopy. For transmission electron microscopy, the worms were fixed in 0.1M phosphate-buffered 2.5% glutaraldehyde, post-fixed for 1 hr in 1% osmium tetroxyde in the same buffer, dehydrated in an increasing graded ethanol series, and embedded in Epon (Luft 1961). Ultrathin sections were collected on copper grids, double-stained with 2% alcoholic uranyl acetate and lead citrate (Reynolds 1963) and examined in a Zeiss EM 900 electron microscope. RESULTS The morphology of M. macracantha by light and scanning electron microscopy was recently published (Kohn et al. 1994). The testes of M. macracantha is composed of numerous follicles, each of which includes different stages of cellular development of spermatogenesis. The primary spermatogonia produce primary spermatocytes by mitotic divisions. The secondary spermatocytes are formed by the same divisions. By meiosis, numerous spermatids originate from the spermatocytes and each one differentiates into a mature spermatozoon. Spermatogonia -The spermatogonia appear as irregularly-shaped cells (Fig. 1). By mitotic divisions, they divide twice, forming secondary and tertiary spermatogonial stages. The nucleus of these cells is almost spherical, located centrally in the cells and occupies most of the cell volume. It contains a single nucleolus and irregular clumps of heterochromatin scattered throughout a granular nucleoplasm. Few ribosomes are attached to the nuclear envelope. The cytoplasm of the spermatogonia contains few membranous elements. Many free ribosomes were observed as clusters or rosettes (polyribosomes). Small mitochondria have short cristae and weakly stained matrices. In the primary and secondary spermatogonia, the mitochondria are located at the cell periphery. In the tertiary stage the nucleus becomes displaced to the border of the cell, and the mitochondria become more numerous and accumulate at the opposite side (Fig. 2). A pair of centrioles near the nucleus was observed (Fig. 3). Spermatocytes - Mitotic divisions of the tertiary spermatogonia results in the production of eight spermatocytes, joined at their bases around the central cell portion. The spermatocytes are the largest cells in the process of spermatogenesis. The nucleus is proportionally smaller than that of the spermatogonia, with granular nucleoplasm containing sparse and condensed chromatin (Fig. 4). Synaptonemal complexes were observed within the chromatin mass of the primary spermatocytes, indicating the pairing of homologous chromosomes during meiosis (Fig. 5). The spermatocytes possess a rough endoplasmatic reticulum (Fig. 9), Golgi cisterns (Fig. 6), and numerous free ribosomes inside the cytoplasm. The mitochondria are similar to those of spermatogonia. Spermatids - The spermatids (Fig. 8) are formed by meiotic cell division of secondary spermatocytes. Each spermatid differentiates into a mature spermatozoon. The nuclei of the spermatids are smaller than those of the spermatocytes. The cytoplasm contains mitochondria, endoplasmatic reticulum and groups of free ribosomes. Spermiogenesis - Spermiogenesis is characterized by the differentiation of the spermatids to spermatozoa. A group of spermatids fuse forming an irregular cytoplasmatic mass; all the spermatids of the same mass are at the same developmental stage (Fig. 8). This cytoplasmatic mass produces elongated projections, known as the 'zone of differentiation' (Fig. 12). The spermatid nuclei become elongated, moving into the cytoplasmatic projections, close to the plasma membrane, which is supported by a single row of microtubules. Basal bodies and associated rootlet fibers (Fig. 7), intercentriolar bodies, numerous free ribosomes, and rough endoplasmatic reticulum, were also observed in the zone of differentiation. After the flagellar elements have rejoined the main central process, and the nucleus and mitochondria have migrated into the projection, the zone of differentiation constricts and each spermatozo-on is released from the general cell mass (Fig. 11). Spermatozoa - The immature spermatozoa present a nucleus of low electron density, mitochondria not yet fused, axonemes and cortical peripheral microtubules (Fig. 13). The mature spermatozoon is filiform, and can be divided in three regions: anterior, middle and terminal. Observations on longitudinal sections show an elongated nucleus from the anterior to the middle region of the spermatozoa, and a single mitochondrion situated parallel to the nucleus (Fig. 10). Cross sections of the anterior and middle regions of mature spermatozoa show the nucleus with strongly condensed chromatin, the mitochondrion with short cristae and two axonemes (Fig. 10). Each axoneme presents a central unit surrounded by nine pairs of microtubules (9 + 1). The plasma membrane is supported by a single layer of microtubules, disposed parallel to the longitudinal axis of the spermatozoa (Fig. 15). The number of peripheral microtubules reduces in the middle region (Fig. 14). Thereafter this layer of peripheral microtubules is reorganized in two rows: one opposite and the other near to the mitochondrion. The axonemes remain joined at the proximal part of the terminal region (Fig. 16), and separate in two flagella at the distal part (Fig. 17). DISCUSSION Recently, Justine published the bibliographical data about the ultrastructure of monogeneans by scanning and transmission electron microscope observations and about spermiogenesis and spermatozoa of monogeneans (Justine 1993a,b). Studies of spermiogenesis and spermatozoa on Microcotylidae had been published by several authors (Tuzet & Ktari 1971a,b, Yousif & Crane 1979, Justine & Mattei 1985, Schmahl & Obiekezie 1991, Justine 1992, Rohde & Watson 1994a,b). The ultrastructural arrangement of the testis and the cytological changes during spermiogenesis in M. macracantha are in agreement with the patterns already established for the Monogenea. Similar to other species of Oligonchoinea, as in Diclidophora merlangi (see Halton & Hardcastle 1976), Protomicrocotyle ivoriensis and Gastrocotyle sp. (see Schmal & Obiekezie 1991), spermatogonia in M. macracantha present the mitochondria and the free ribosomes as the main organelles found in the cytoplasm. The Golgi complex occupies a typical position within the spermatocyts; its trans-face is directed to the plasma membrane. This pattern was also observed by Halton and Hardcastle (1976) in D. merlangi. The presence of two axonemes with the 9 + 1 configuration, the nucleus, mitochondria and cortical longitudinal microtubules can be observed at transversal sections and constitutes a typical pattern for the majority of parasites of this subclass. The cortical microtubules are absent in almost all Polyonchoinea, whereas in Oligonchoinea, they occur in most of the species studied (Rohde 1971, 1980, Tuzet & Ktari 1971b, Halton & Hardcastle 1976, Justine & Mattei 1984), except in D. gracile which present aflagellate spermatozoa (Justine et al. 1985a). The cortical microtubules of M. macracantha may be considered as reinforcement elements which develop at the beginning of the transformation process. Justine et al. (1985b) have proposed the use of spermatozoa ultrastructure for phylogeny studies of Monogenea; they use two spermatozoan characters to classify the group: the number of axonemes and the presence or absence of cortical microtubules.The spermatozoa of M. macracantha clearly belongs to the type 1 referred by these authors.
ACKNOWLEDGEMENTS To Prof. DW Halton from The Queen's University, Belfast, U.K., for reading the manuscript; to Dr Marilia Taffarel from Instituto Oswaldo Cruz and to the referee of this paper, for the suggestions. REFERENCES Halton DW, Hardcastle A 1976. Spermatogenesis in a monogenean, Diclidophora merlangi. Internat J Parasitol 6: 43-53. Justine JL 1991a. Cladistic study in the Monogenea (Platyhelminthes), based upon a parsimony analysis of spermiogenetic and spermatozoal ultrastructural characters. Internat J Parasitol 21: 821-838. Justine JL 1991b. Phylogeny of parasitic Platyhelminthes: a critical study of synapomorphies proposed on the basis of the ultrastructure of spermiogenesis and spermatozoa. Can J Zool 69: 1421-1440. Justine JL 1992. Ultrastructure of spermiogenesis, spermatozoa and the tegument in Astriaster sp. (Platyhelminthes, Monogenea, Polyopisthocotylea, Microcotylidae). Zool Scripta 21: 231-238. Justine JL 1993a. Phylogenie des Monogenes basee sur une analyse de parcimonie des caracteres de l'ultrastructure de la spermiogenese et des spermatozoides incluant les resultats recents. Bull Fr PEche Piscic 328: 137-155. Justine JL 1993b. Ultrastructure des Monogenes listes des especes et des organes etudies. Bull Fr PEche Piscic 328: 156-188. Justine JL, Mattei X 1984. Ultrastructure du spermatozoide du Monogene Hexostoma (Polyopisthocotylea, Hexostomatidae). Ann Parasit Hum Com 59: 227-229. Justine JL, Mattei X 1985. Particularites ultrastructurales des spermatozoides de quelques Monogenes Polyopisthocotylea. Ann Sci Nat Zool 7: 143-152. Justine JL , Brun L, Mattei X 1985a. First report of aflagellate spermatozoon in a parasitic Platyhelminthe, found in the monogenean Diplozoon gracile (Polyopisthocotylea, Diplozoidae). Ann Parasit Hum Comp 60: 761-762. Justine JL, Lambert A, Mattei X 1985b. Spermatozoon ultrastructure and phylogenetic relationships in the monogeneans (Platyhelminthes). Internat J Parasitol 15: 601-608. Kohn A, Cohen SC, Baptista-Farias MFD 1994. A redescription of the morphology of Metamicrocotyla macracantha (Alexander, 1954) Koratha, 1955 (Monogenea, Microcotylidae) from Mugil liza in Brazil. Syst Parasit 27: 127-132. Lambert A 1980. Oncomiracidiums et phylogenese des Monogenea (Plathelminthes) deuxieme partie: phylogenese des Monogenea. Ann Parasit Hum Com 55: 281-325. Lebedev BI 1988. Monogenea in the light of new evidence and their position among platyhelminthes. Ang Parasit 29: 149-167. Luft JH 1961. Improvements in epoxi resin embedding-methods. J Biophy Cytol 9: 409. Malmberg G 1990. On the ontogeny of the haptor and the evolution of the Monogenea. Syst Parasit 17: 1- 65. Reynolds ES 1963. The use of lead citrate at high pH as electron opaque stain in electron microscopy. J Cell Biol 17: 208. Rohde K 1971. Phylogenetic origin of trematodes. Parasit Schrif 21: 17-27. Rohde K 1980. Some aspects of the ultrastructure of Gotocotyla secunda and Hexostoma euthynni. Ang Parasit 21: 32-48. Rohde K, Watson NA 1994a. Spermiogenesis in Gonoplasius sp. (Monogenea, Polyopisthocotylea, Microcotylidae). Acta Parasitologica 39: 111-116. Rohde K, Watson NA 1994b. Ultrastructure of spermiogenesis and spermatozoa of Polylabroides australis (Platyhelminthes, Monogenea, Polyopisthocotylea, Microcotylidae). Parasite 1: 115-122. Schmall G, Obiekezie A 1991. Fine structure of spermatogenesis in polyopisthocotylid monogeneans Protomicrocotyle ivoriensis (Gastrocotyle sp.). Parasit Res 77: 115-122. Tuzet O, Ktari MH 1971a. La spermiogenese et la structure du spermatozoide de Microcotyle mormyri Lorenz, 1878 (Monogenea). Comptes Rendus de lÆAcademie des Sciences (Paris) serie D Sci Natur 272: 2702-2705. Tuzet O, Ktari MH l971b. Recherches sur lÆultrastructure du spermatozoide de quelques Monogenes. Bull Soc Zool France 96: 535-540. Yousif MS, Crane JW l979. Ultrastructure and spermiogenesis Microcotyle sebastis (Goto, 1894) (Monogenea), p. 11-13. In G Bylund, Proc IX Symp Scandinavian Soc Parasitol Abo Finland. This paper is part of the thesis of Master Degree of the first author. +Research fellows "Conselho Nacional de Desenvol-vimento Cientifico e Tecnologico - CNPq". Received 6 July 1994 Accepted 10 November 1994
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95120d.gif] [oc95120e.gif] [oc95120c.gif] [oc95120b.gif] [oc95120a.gif]Photo images[oc95120d.jpg] [oc95120a.jpg] [oc95120b.jpg] [oc95120e.jpg] [oc95120c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}