 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Larval and Pupal Periods of Peckia chrysostoma and Adiscochaeta ingens (Diptera: Sarcophagidae) Reared under Laboratory Conditions Marisa Vianna Ferraz Departamento de Entomologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
Code Number: OC95122
Size of Files:
Text: 14K
Graphics: Line Drawings (gif) 50K
Peckia chrysostoma obtained mean viability of 97.0+/-
2.4% for larvae and of 96.9+/-2.5% for pupae (total viability
of 94.0+/-3.7%). Adiscochaeta ingens obtained mean
viability of 93.0+/-7.5% for larvae and of 92.8+/-7.6% for
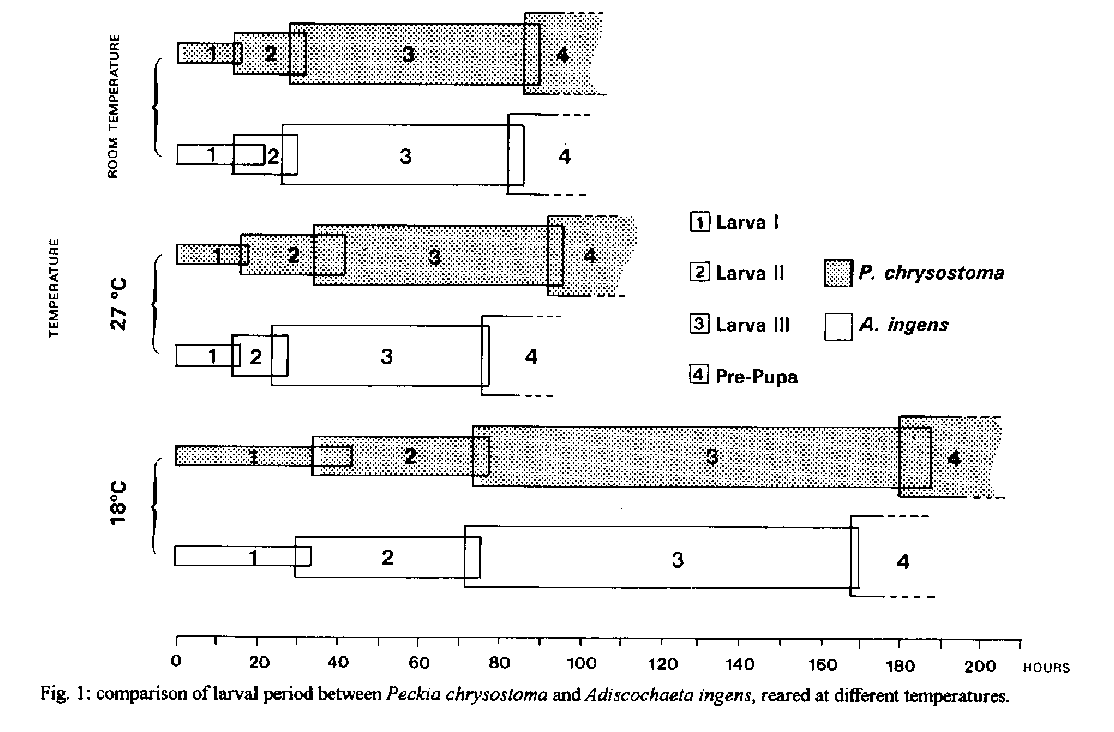
pupae (total viability of 86.0+/-7.3%).P. chrysostoma obtained mean larval period of 185+/-4 hr at 18 C, of 94+/-2 hr at 27 C and of 88+/-2 hr at room temperature (range of 23 C and 29 C). A. ingens obtained mean larval period of 169+/-1 hr at 18 C, of 77+/-1 hr at 27 C and of 84+/-2 hr at room temperature.
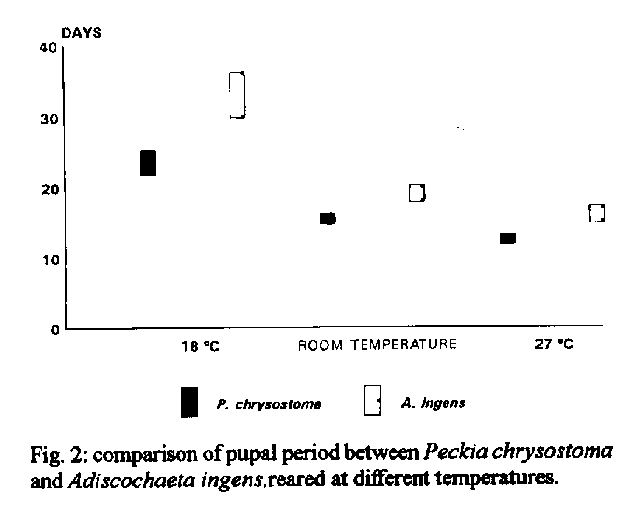
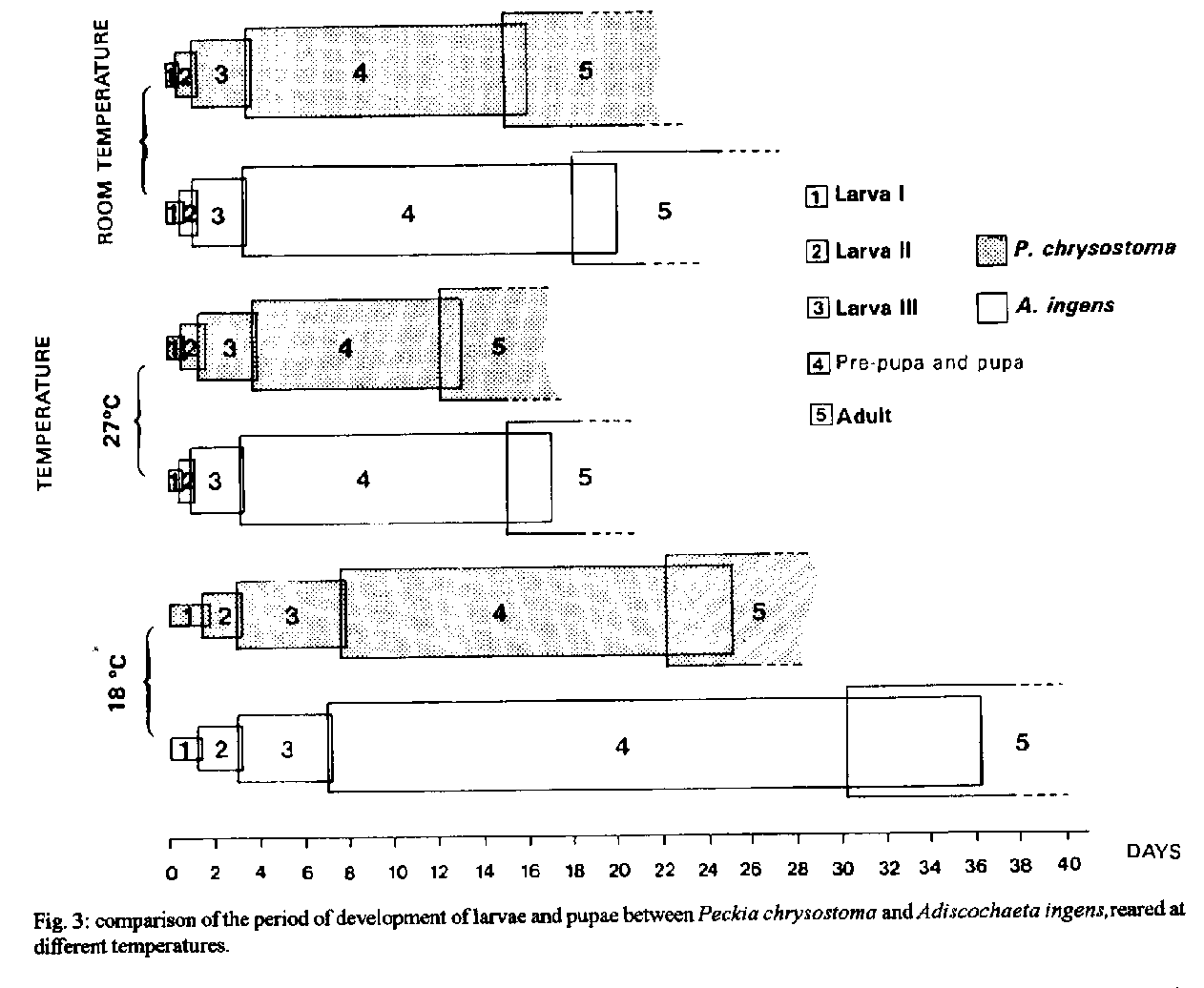
P. chrysostoma obtained mean pupal period of 23.5+/-1.3 days at 18 C, of 12.5+/-0.7 days at 27 C and of 15.5+/-0.7 days at room temperature. A. ingens obtained mean pupal period of 33.0+/-2.2 days at 18 C, of 16.0+/-1.0 days at 27 C and of 19.0+/-1.0 days at room temperature. Key words: larval/pupal period - larval/pupal viability - Peckia chrysostoma - Adiscochaeta ingens - Sarcophagidae - Diptera This study is part of a series (Ferraz 1992a, b, 1993, 1994, among others), carried out at the laboratory, on the comparative biology of Peckia chrysostoma (Wiedemann 1830) and Adiscochaeta ingens (Walker 1849), two caliptratae muscoids. Results shown here compare the larval and pupal period and viability between one autochthonous species, P. chrysostoma, and an introduced one, A. ingens. P. chrysostoma, a neotropic and sinantropic species, is widely spread and frequently found in high densities in Rio de Janeiro (d'Almeida 1984, Tavares et al. 1988, Ferraz 1992a, b, 1993). A. ingens, originally from the Brazilian 'campos cerrados', has been found in Rio de Janeiro, but in numbers of little significance (Lopes 1973, 1974, Linhares 1981, Lopes & Tibana 1982, d'Almeida 1984). MATERIALS AND METHODS The methodology adopted for the field collection, colonization and rearing of adults has been previously described (Ferraz 1993). Larval viability - Five groups of 20 recently laid larvae were maintained in containers with rearing media. The percentage of formed pupae was observed daily. This experiment was carried out at room temperature. Larval period at different temperatures - Six containers with rearing media in the proportion of 2g per larva were prepared. Each container had 65 recently laid larvae (totaling approximately 400 larvae) which were maintained at 27 C at 70-80% relative humidity (R.H.) (B.O.D. refrigerators). Every 2 hr, 15 larvae were taken to determine the larval instar by observing the respiratory spiracles. We did not take the last 15 larvae of each container for this purpose as low density of individuals may alter the time of development. Comparisons were made with groups of the same number maintained at 18 C at 70-80% R.H. (B.O.D. refrigerators) and at room temperature (control-groups). Pupal viability - Five groups of 20 pre-pupae were kept in containers with moist sawdust. The percentage of emerged adults was observed daily. This experiment was carried out at room temperature. Pupal period at different temperatures - The pre-pupal period begins when the larva leaves the culture media, hides in the moist sawdust and moves up to an appropriate local. There, it stays still, retracts its extremities and begins to darken its tegument, transforming itself into a pupa. Here we considered the pupal period as including the pre-pupal period. Three groups of 20 pre-pupae were put into a container with moist sawdust and maintained at 27 C at 70-80% R.H. (B.O.D. refrigerator). Every 24 hr they were observed to determine the pupal period. Comparisons were made with groups of the same number maintained at 18 C at 70-80% R.H. (B.O.D. refrigerator) and at room teperature (control-groups). The experiments conducted at room temperature had mean temperature of 25.9 C (from 23 C to 29 C) and 76% mean R.H. (59% - 88%). RESULTS Table I displays the higher larval and pupal viability of P. chrysostoma and A. ingens. Only viability of the pupae was significantly different between the two species (Table III). Viability of the larvae was not. Furthermore, there was no significant difference between the viability of larvae and pupae for P. chrysostoma and A. ingens (Table II). Fig. 1 displays the larval period observed at each temperature, for each stage, every two hours, of P. chrysostoma and A. ingens, showing the faster larval period of A. ingens. Fig. 2 displays the pupal period observed at each temperature, once a day, of P. chrysostoma and A. ingens, showing the faster pupal period of P. chrysostoma. Fig. 3 displays the total immature stages of development demonstrating the faster development of P. chrysostoma at the three conditions studied and the faster development of both species at 27 C compared to room temperature and to 18 C.
TABLE 1 Larval and pupil viability of Peckia chrysostoma and Adiscochaeta ingens at room temperature
-------------------------------------------------------
P. chrysostoma A.ingens
-------------------------------------------------------
Initial larvae No. 100 100
L 97 93
P 94 86
V 95.5 89.5
-------------------------------------------------------
L: number of formed pupae (larval viability); P: number of emerged adults (pupil viability); V: mean total viability - from larva to adult TABLE II Chi-square analysis of viability of Peckia chrysostoma and Adiscochaeta ingens in relation to the immature stages. Number in parentheses indicate the expected values according to the null hypothesis.
------------------------------------------------------------
Immature stages
Species Viability Larvae Pupae Statistics
------------------------------------------------------------
Pc yes 97 (95.5) 94 (95.5) X^2 = 1.047
no 3 (4.5) 6 (4.5) p > 0.05
Ai yes 93 (89.5) 86 (89.5) X^2 = 2.607
no 7 (10.5) 14 (10.5) p > 0.05
------------------------------------------------------------
X^2: chi-square; p: probability; Pc: Peckia chrysostoma; Ai:
Adiscochaeta ingens. Degree of freedom = 1
Fig. 2: comparison of pupal period between Peckia chrysostoma and Adiscochaeta ingens reared at different temperatures. Fig. 3: comparison of the period of development of larvae and pupae between Peckia chrysostoma and Adiscochaeta ingens reared at different temperatures.
------------------------------------------------------------
Immature Species
stages Viability Pc Ai Statistics
------------------------------------------------------------
Larvae yes 97(95.0) 93(95.0) X^2 = 1.684
no 3(5.0) 7(5.0) p > 0.05
Pupae yes 94(90.0) 86(90.0) X^2 = 3.556
no 6(10.0) 14(10.0) p< 0.05
-------------------------------------------------------------
X^2: chi-square; p:probability; Pc; Peckia chrysostoma
Ai: Adiscochaeta ingens. Degree of freedom = 1DISCUSSION Several authors have already observed that viability of immature stages varies according to the species. Viability of larvae and pupae here was relatively high for both species. Despite the higher viability of P. chrysostoma, it was not significantly different, for the larvae, than that of A. ingens. For Sarcophaga cooleyi, S. shermani and S. bullata (Kamal 1958) the viabilities of the pupae was of 74-91%, 81-93% and 64-87%, respectively. S. haemorrhoidalis (Madubunyl, 1986) obtained viability of 80.69+/-3.20 for the larvae and 89.83+/- 3.80 for the pupae. The time of development of muscoid diptera differs greatly according to the species. For S. cooleyi, S. shermani and S. bullata, Kamal (1958) also found differences: an average of 24, 22 and 26 hr, respectively, for the development of the first larval instar; 18, 16 and 18 hr for the second larval instar; 48, 48 and 54 hr for the third larval instar; 96, 104 and 112 hr for the pre-pupa stage; and 9, 8 and 12 hr for the pupal period. For S. haemorrhoidalis (Madubunyl, 1986) the pupal period lasted four days and, from the egg to the adult, lasted approximately 32 days (= 768 hr). As expected here, both species developed faster in a higher temperature (27 C) than in a low one (18 C). Comparing with these results, Sutherland (1979, 1980) observed that pupae of Stomoxys calcitrans bore temperatures between 20 C and 30 C and the larvae prefered temperatures between 19.5 C and 32.5 C. Cook et al. (1980) and Cook and Spain (1982) observed that Haematobia irritans pupae bore temperatures between 17.5 and 35 C. Survival of larvae and pupae was optimized at 25 C. Excluding the P. chrystosoma larvae, the life cycle of larvae and pupae for both species was faster under constant and controlled higher temperatures than under variable conditions. Kamal, in 1958, observed the same: constant and controlled temperatures speeded the life cycle but shortened the lifespan of adults of some calliphorid and sarcophagid, compared to results obtained under variable temperature conditions. ACKNOWLEDGEMENTS To Dr Hugo de Souza Lopes, for his inestimable supervision during this study. To Dr Rubens Pinto de Mello, for his valuable observations and comments. To Dr Otavio Pieri, for the statistical analisys. To Denise Tavares Goncalves, for the English review. REFERENCES Cook IM, Spain AV 1982. The effect of temperature and moisture on survival of the immature stages of the buffalo fly, Haematobia irritans exigua de Meijere (Diptera: Muscidae). Aust J Zool 30: 923-930. Cook IM, Spain AV, Sinclair DF 1980. The effects of the temperature and moisture levels of the larval breeding medium on puparial form in the buffalo fly, Haematobia irritans exigua (Diptera: Muscidae). Aust J Zool 28: 547-552. d'Almeida JM 1984. Sinantropia de Sarcophagidae (Diptera) na regiao metropolitana do Estado do Rio de Janeiro. Arq Univ Fed Rur Rio de Janeiro 7: 101-110. Ferraz MV 1992a. Estudos comparados do desenvolvimento biologico e do comportamento de Peckia chrysostoma (Wiedemann, 1830) e Adiscochaeta ingens (Walker, 1849) (Diptera: Sarcophagidae) em condicoes de laboratorio. Msc. Thesis. Instituto Oswaldo Cruz, FIOCRUZ, Rio de Janeiro, Brazil, 183 pp. Ferraz MV 1992b. Comparison of the reproductive behavior between isolated Peckia chrysostoma (Wiedemann, 1830) and Adiscochaeta ingens (Walker, 1849) (Diptera: Sarcophagidae) females reared in laboratory. Mem Inst Oswaldo Cruz 87: 131-139. Ferraz MV 1993. Interespecific competition between Peckia chrysostoma and Adiscochaeta ingens, (Diptera: Sarcophagidae) larvae reared in laboratory. Mem Inst Oswaldo Cruz 88: 189-194. Ferraz MV 1994. Longevity of adults of Peckia chrysostoma and Adiscochaeta ingens (Diptera: Sarcophagidae) reared with or without protein. Mem Inst Oswaldo Cruz 89: 421-424. Kamal AS 1958. Comparative study of thirteen species of sarcosaprophagous Calliphoridae and Sarcophagidae (Diptera) I. Bionomics. Ann Ent Soc Am 51: 261-271. Linhares AX 1981. Synanthropy of Calliphoridae and Sarcophagidae (Diptera) in the city of Campinas, Sao Paulo, Brazil. Rev brasil Entomol 25: 189-215. Lopes HS 1973. Collecting and rearing Sarcophagid flies (Diptera) in Brazil, during forty years. An Acad brasil Cienc 45: 279-291. Lopes HS 1974. Sarcophagid flies Diptera from Pacatuba, State of Ceara, Brazil. Rev brasil Biol 34: 271-294. Lopes HS, Tibana R 1982. Sarcophagid flies (Diptera) from Sinop, State of Mato Grosso, Brazil. Mem Inst Oswaldo Cruz 77: 285-298. Madubunyl LC 1986. Laboratory life history parameters of the red-tailed fleshfly, Sarcophaga haemorrhoidalis (Fallen) (Diptera: Sarcophagidae). Insect Sci Appl 7: 617-621. Sutherland B 1979. The effects of temperature on the adults, eggs and pupae of Stomoxys calcitrans Linnaeus (Diptera: Muscidae). Onderstepoort J Vet Res 46: 223-227. Sutherland B 1980. The temperature preferences of the motile stages of Stomoxys calcitrans Linnaeus (Diptera: Muscidae). Onderstepoort J Vet Res 47: 7-11. Tavares DES, Porthun JR, Fernandes EA, Oliveira SP 1988. Estudo sobre comunidades de sarcofagideos sinantropicos (Diptera, Sarcophagidae) no Instituto Oswaldo Cruz, Manguinhos, RJ. p.233. In Res XV Congr Brasil Zool, Curitiba, PR.
Received 14 December 1994 Accepted 8 May 1995 Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc95122b.gif] [oc95122a.gif] [oc95122c.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}