 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Infection of Oxydoras kneri Bleecker, 1862 (Pisces, Doradidae) by the Acanthocephalan Paracavisoma impudica (Diesing, 1851) Kritcher, 1957
JC Eiras, GC Pavanelli*, MH Machado* Instituto de Zoologia, Faculdade de CiEncias, Universidade do Porto, 4050 Porto, Portugal *Departamento de Biologia/Nupelia, Fundacao Universidade Estadual de Maringa, Maringa, PR, Brasil
Code Number: OC95127
Size of Files:
Text: 9K
Graphics: Photos (jpg) 93K / Halftones (gif) 158K
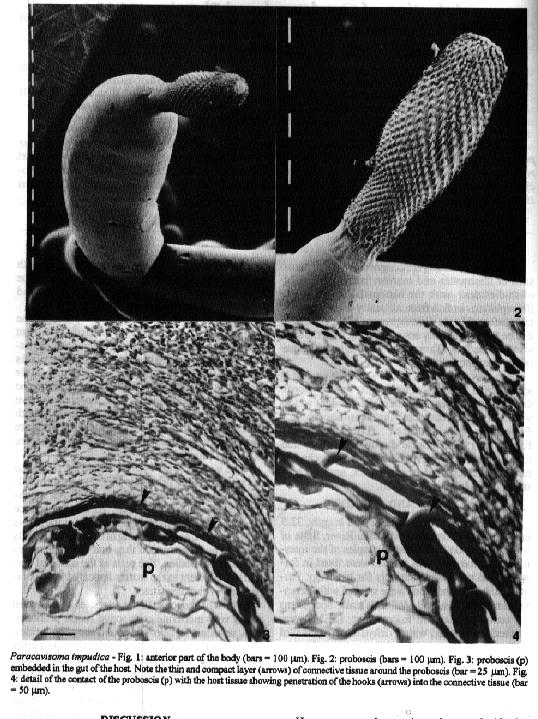
Infection of Oxydoras kneri by the acanthocephalan Paracavisoma impudica is described. The parasites do not penetrate deeply into the host gut wall and do not reach the muscularis layers. Host reaction is minimal, consisting of limited fibrosis around the proboscides. Haemorrhages and lymphocyte infiltration are not observed, and phagocytic cells are only present occasionally. Oxydoras kneri is a newly reported host for Paracavisoma impudica. Key words: Oxydoras kneri - Paracavisoma impudica - fishes - acanthocephala - histopathology - Brazil Acanthocephalans are common intestinal parasites of freshwater and marine fishes. A number of studies deal with the histopathology of acanthocephalans in fish from naturally or experimentally infected hosts. Consequences of the infection can be few, apparently not interfering with the survival of the host (Taraschewski 1989), or a cause of the host mortality (Jilek 1979). Recently, one of us (GC Pavanelli) caught a specimen of the tropical freshwater fish Oxydoras kneri (Siluriformes, Siluroidei, Doradidae) which was heavily infected by the acanthocephalan Paracavisoma impudica (Diesing, 1851) Kritecher, 1957. Because the histopathology due to this parasite has never been studied, its characteristics are reported in this paper, and the host status of O. kneri is discussed. MATERIALS AND METHODS The specimen of O. kneri (total length: 32.5 cm) was fished from the Miranda River, State of Mato Grosso do Sul, Brazil. Samples of the intestine, with the parasites in situ, were fixed in formalin and processed routinely for histology. Paraffin embedded sections were stained with Hematoxylin and Eosin, and MalloryÆs stain. Specimens for scanning electron microscopy (SEM) were washed in distilled water, dehydrated in ethanol series, critical-point-dried in carbon dioxide, coated with gold/palladium and observed in a Phillips 501 SEM at 7.2 kV. RESULTS Observation of the intestine of the host showed it was heavily infected, the parasites being scattered throughout the length of the intestine, with up to 25 parasites per cm^2 in some places. The worms were 0.7-0.9 mm in total length, and gravid females were not observed. The wall of the intestine is composed of the inner epithelium, lamina propria, stratum granulosum surrounded by a loose connective tissue layer, muscularis circularis, muscularis longitudinalis and serosa. The muscularis circularis is about 3.5 times thicker than the muscularis longitudinalis. The loose connective tissue layer is the thickest, being thicker than both the two muscularis layers. The intestinal folds are composed of a columnar epithelium and a thin core of lamina propria.
The worms (Fig. 1) were weakly attached to the intestine, the proboscis (Fig. 2) being embedded in the loose connective tissue but not reaching the muscularis circularis. As a consequence there was destruction of the epithelium at the points of penetration, and deepening of the intestinal folds near the worm body, as if they had been pushed by the parasites bodies. The cells of the mucosal epithelium adjacent to the worms appeared to be compressed by the parasites. The host response to the parasites consisted in just a very moderate connective tissue proliferation, resulting in fibrosis around the proboscides, the connective fibers forming a thin compact layer about 20 mm thick (Fig. 3). This layer had occasional ruptures especially in places which were in contact with the proboscis hooks (Fig. 4). Along the penetration points a small number of necrotic cells was observed, and sometimes cellular debris was attached to the parasites proboscis. Occasional macrophages were present in the tissues surrounding the proboscides, while haemorrhages or lymphocyte infiltration were not observed. DISCUSSION Paracavisoma impudica was described from the host Pseudodoras niger (Doradidae). The present paper reports for the first time its occurrence in O. kneri. However, our observations do not clarify the host status of O. kneri. Some acanthocephalan species incorporate an obligate or accidental paratenic host into the life cycle. The paratenic hosts can became infected accidentaly by predation on fish with immature worms. They will develop further if the paratenic host is eaten by the definitive one. On the other hand, a paratenic host can be ecologically essential in case the definitive host does not predate the intermediate one. Due to the absence of gravid females from our host it is difficult to state wether O. kneri is a final or a paratenic host. Histopathology in the fish intestine caused by acanthocephalans is primarily related to the depth of proboscis penetration (Bullock 1963). Complete penetration of the gut wall was described for a number of species from different hosts (Taraschewski 1988, 1989, Wanstall et al. 1988). On the other hand, a number of histopathological features such as fibrosis, haemorrhages, inflammation, cell necrosis, etc., were reported for several species (Taraschewski 1988, 1989, Wanstall et al. 1988). In our material the parasites did not penetrate deeply into the wall of the intestine, and did not reach the first muscular layer. Therefore, the damage to the host gut seems limited to the mucosa and submucosa layers, probably causing some degree of impairment of the absorption power of the affected area. Consequently, the nutritional state of the host is likely to be decreased and, as the mucosa is damaged, there is most probably some alteration in the osmoregulatory process. On the other hand, the wounds produced by the parasites can expose the host to secondary infections by pathogenic microorganisms. This is especially important in the case where the worms can change their attachment sites, therefore producing multiple wounds. The present observations did not demonstrate whether the parasites can change their site of attachment. However, due to the limited fibrosis produced around the proboscides, that possibility seems likely. The histopathological findings observed lead us to the conclusion that the parasite-induced pathogeny caused by each individual parasite is not severe, apparently not having a marked influence upon the host metabolism. However, in the case of heavy infections, as in this report, it is presumed that serious consequences can occur due to the cumulative lesions produced by the parasites, especially if the host is small in size. Moreover, it is probable that heavy infections by the parasite might cause blockage of the lumen of the gut. As stated by Wanstall et al. (1988), occlusion of the hostÆs gut is not an uncommon consequence of acanthocephalan infection in fish. ACKNOWLEDGEMENTS To Dr CJ Veltkamp, University of Liverpool, for facilities provided for examining the parasites under scanning electron microscopy. REFERENCES Bullock WL 1963. Intestinal histology of some salmonid fishes with particular reference to the histopathology of acanthocephalan infections. J Morphol 112: 23-44. Jilek R 1979. Histopathology due to the presence of Gracilisentis gracilisentis in Dorosoma cepedianum (Le Sueur). J Fish Biol 14: 593-595. Taraschewski H 1988. Host-parasite interface of fish acanthocephalans. I. Acanthocephalus anguillae (Palaeacan-thocephala) in naturally infected fishes: LM and TEM investigations. Dis aquat Org 4: 109-119. Taraschewski H 1989. Host-parasite interface of Paratenuisentis ambiguus (Eoacanthocephala) in naturally infected eel and in laboratory-infected sticklebacks and juvenile carp and rainbow trout. J Parasitol 75: 911-919. Wanstall ST, Thomas JS, Robotham PWJ 1988. The pathology caused by Pomphorhynchus laevis Muller in the alimentary tract of the stone loach, Noemacheilus barbatulus (L.). J Fish Diseases 11: 511-523.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Halftone images[oc95127a.gif]Photo images[oc95127a.jpg] |
| |||||||||

{kind=link}