 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Eurytrema coelomaticum: Influence of the Infection on the Reproduction and Nucleic Acids Contents in the Albumen Gland and Ovotestis of Bradybaena similaris Jairo Pinheiro, Suzana B Amato* Departamento de CiEncias Fisiologicas *Departamento de Biologia Animal, Universidade Federal Rural do Rio de Janeiro, Caixa Postal 74.512, 23851-970 Seropedica, RJ, Brasil
Code Number: OC95129
Size of Files:
Text: 18K
Graphics: Line Drawings (gif) 12K
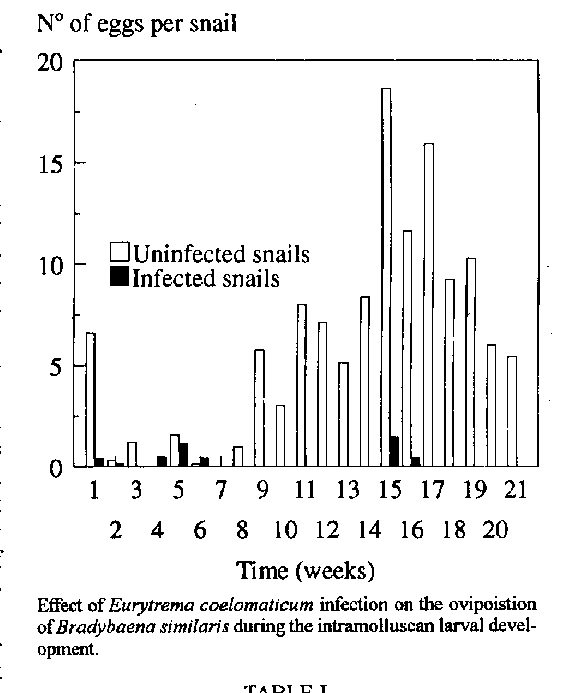
The number of eggs laid per snail in Bradybaena similaris and the nucleic acids (DNA and RNA) in the albumen gland and ovotestis were quantified in snails infected with sporocysts of the digenetic trematode Eurytrema coelomaticum. The total number of eggs laid per mollusc was reduced by 96.32% at the end of the larval development. The DNA concentration increased by 700%, and the RNA concentration was reduced by 8.38% by the time when the daughter sporocysts of E. coelomaticum were released from B. similaris. The relation between these values and the inhibition of the reproduction observed in infected molluscs is discussed. Key words: Eurytrema coelomaticum - Bradybaena similaris - parasitic castration - nucleic acid - ovotestis - albumen gland The trematode Eurytrema coelomaticum (Giard et Billet, 1892) Looss, 1907 (Digenea, Dicrocoeliidae) has a complex life cycle with two intermediate hosts. In Brazil, snails of the species Bradybaena similaris (Ferussac, 1821) are the first intermediate host and orthopteran tettigoniids of the genus Conocephalus Thunberg, 1815, are the second intermediate host (Mattos Jr et al. 1987). A consequence of the snail infection by larval digenetic trematodes is the interruption of the normal activities of reproduction in its snail hosts, parasitic castration (Baudoin 1975). This process has been observed and studied in several host/parasite interfaces (Brumpt 1941, Pan 1965, Sturrock 1966, McClelland & Bourns 1969, Sturrock & Sturrock 1970, Webe & James 1972, Loker 1979, Wilson & Denison 1980, Sullivan et al. 1985). Paschoal (1991), using histology, observed degeneration of the ovotestis in snails, B. similaris, infected with sporocysts of E. coelomaticum, with reduction in the spermatogenesis. This author also noted a reduction in the snail's oviposition. Our objective was to study the effect of parasitism by larvae of E. coelomaticum on the reproductive function of its snail host, the snail's oviposition; the DNA and RNA content of the albumen gland and ovotestis were quantified in uninfected and infected snails. MATERIALS AND METHODS Maintenance and snail infection - Specimens of B. similaris were collected from horticultural plots at km 45, of the Antiga Rodovia Rio de Janeiro-Sao Paulo (BR 465), Itaguai, RJ, Brazil. The snails were mantained in vivaria under laboratory conditions (25 +/- 3 C; 80 +/- 7% relative humidity). All snails were examined through their transparent shells for the presence of Postharmostomum gallinum metacercariae in the pericardial cavity. Samples of randomly chosen snails were dissected to ensure that the snails were free of larval helminths. Snails free of infection were maintained in glass vivaria with earth at the bottom and moistened with tap water at two day intervals. They were fed ad libitum with cabbage leaves (Brassica sp.) and ration elaborated according to Frantz and Mossman (1989). Eggs of E. coelomaticum were obtained from adult worms collected from the pancreas of naturally infected cattle slaughtered at the Matadouro Industrial de Santa Cruz, Rio de Janeiro, RJ, Brazil. The worms were placed in petri dishes with 0.85% saline, and left for 12 hr at room temperature (20 +/- 5 C) in order to lay eggs. The eggs were selected according to Tang (1950), washed in distilled water and pipetted over pieces of cabbage leaves placed in petri dishes lined with filter paper moistened with tap water. Five or ten snails, with 10 mm of shell diameter were placed on these leaves, in a proportion of 10 eggs/snail, and left overnight. The next day they were transferred to the vivarium. Snail's oviposition - Weekly, the earth of each vivarium was carefully examined for the eggs of B. similaris, in infected and uninfected snails. The number of eggs produced was recorded and related to the number of snails of each vivarium. Quantitative evaluation of nucleic acids in the albumen gland-ovotestis complex - The snail shells were broken, and the snails individually dissected under 0.65% saline. The albumen gland-ovotestis complex was separated from the seminal vesicle and the other parts. Snails were dissected at 30, 60, 90, and 150 days post-infection. Control uninfected snails underwent the same procedure. The nucleic acids were extracted by the method of Schimdt and Thannhauser (1945), using tissue from 25 snails for each quantification. DNA and RNA were quantified by the methods of Seibert (1940) and Mejbaum (1939), respectively, in Villela et al. (1977), being expressed in mg of desoxyribose/g of tissue and mg of ribose/g of tissue, respectively. Statistical analysis - The results were submitted to variance analysis using the Student's t-test (a = 5%) . The test of exponential regression was used over the DNA and RNA results (Vieira 1986). RESULTS The snails B. similaris infected with larval E. coelomaticum had a marked reduction on the oviposition (Fig.). A total inhibition of oviposition was observed between the 7th and the 14th, and the 17th and 21st weeks post-infection with E. coelomaticum eggs. The total number of eggs produced during the 150 days of observation was reduced by 96.32% . The result that was significantly different to that observed in the uninfected snails (control group) during the same period (a = 5%).
------------------------------------------------------------
DNA content
(mg desoxyribose/g tissue)
Period of infection Increase in DNA
(days) N X+/-S content (%)
-------------------------------------------------------------
0(control) 5 0.015+/-0.004d 0
30 5 0.020+/-0.008c 33.33
60 5 0.026+/-0.001c 73.33
90 5 0.035+/-0.007b 133.33
150 5 0.120+/-0.014a 700.00
--------------------------------------------------------------
TABLE II. RNA content in the albumen gland-ovotestis complex
of infected and uninfected Bradybaena similaris with
sporocysts of Eurytrema coelomaticum
------------------------------------------------------------
RNA content
(mg desoxyribose/g tissue)
Period of infection Increase in DNA
(days) N X+/-S content (%)
-------------------------------------------------------------
0(control) 5 0.836+/-0.262a 0
30 5 0.293+/-0.035c 64.91
60 5 0.578+/-0.007b 29.94
90 5 0.690+/-0.001a,b 17.65
150 5 0.766+/-0.064a 8.38
--------------------------------------------------------------
X+/- S: mean values +/- standard deviation; N = number of
quantifications; a, b, c and d: means with significant
difference among them (alpha = 5%)
The DNA content in the albumen gland-ovotestis complex of the infected snails progressively increased during the development of the sporocysts (Table I). This increase reached 700% at the end of the larval intramolluscan development. A highly significant difference was observed between the values obtained at the different periods of infection (a = 5%). The RNA content severely decreased at the beginning of the infection, subsequently increasing to values close to normal at the end of the larval development, being only 8.39% lower than the RNA content of the albumen gland-ovotestis complex of the uninfected snails (Table II). Only the RNA content of the albumen gland-ovotestis complex, at the 30th day post-infection, was significantly different from the others (a = 5%). DISCUSSION The results of egg production per snail are similar to the results of McClelland and Bourns (1969), Loker (1979), and Wilson and Denison (1980), and they were used to standardize the data allowing comparisons between uninfected and infected snails. As the snails used during this work were collected from their natural habitat, their age could not be used as parameter. The diameter of the shells was used to evaluate egg production based on a positive correlation observed between snail size and number of eggs produced (Amato & Rosa 1982). The interference of the larval digeneans in the reproduction of their molluscan hosts is a common phenomenon (Wright 1966). According to Cheng et al. (1973) the mechanisms which lead to castration may be associated with mechanical or chemical action of the parasite over the host gonad. Since E. coelomaticum larvae were never found in the gonads of its snail host, B. similaris, the castration is probably due to a secondary effect, which disturbed the host physiology. According to Sullivan et al. (1985), the sporocysts of E. coelomaticum are indirectly interfering with its snail hostÆs reproduction. An answer similar to the one observed in B. similaris infected with E. coelomaticum, in the present study, was registered by McClelland and Bourns (1969) studying the action of Trichobilharzia ocellata over Lymnaea stagnalis reproduction, where the reduction on the egg production by the snails exceeded 95%. The authors concluded that in the interface studied the castration was not due to a direct predation of the gonads by the larvae. The detection of the parasitic castration led us to investigate the mechanism that was involved in this process. A quantification of the nucleic acids was used as parameter to evaluate the rate of cell division in the ovotestis-albumen gland complex of B. similaris infected with E. coelomaticum; Wijdenes et al. (1983) considered the DNA concentration in the albumen gland of L. stagnalis a useful parameter to determine its growth. Studies carried out with T. ocellata/L. stagnalis showed that the maturation of the organs of the reproductive system was arrested in infected snails (Sluiters et al. 1980, Sluiters 1981). This was confirmed by van Elk and Joose (1981) who demonstrated in this same host/parasite interface that the content of DNA did not increase as the snails aged. In B. similaris infected with E. coelomaticum an increase in DNA content was detected in the albumen gland-ovotestis complex during the intramolluscan larval development, indicating that the growth of the albumen gland was not affected by the parasite. Quantification of the RNA content in the albumen gland-ovostestis complex during the larval intramolluscan development of E. coelomaticum was used as indirect means of evaluating protein biosynthesis in these organs and the essential processes for the development of the snail's eggs, egg shell formation and the embryo nutrition. Alterations of this metabolic function would interfere with the normal reproductive activity of the snail. Studying the action of Schistosoma mansoni on the levels of mRNA of the ovotestis and albumen gland of B. glabrata, Crew and Yoshino (1989) observed a reduction in the protein synthesis relating it to a reduction of mRNA. Our results show the same reduction on the RNA content of the albumen gland-ovotestis complex, at the beginning of larval intramolluscan development as well as a subsequent tendency to recover. The content of RNA may be used as a parameter to evaluate the rate of protein biosynthesis, but the inverse may induce an error since in the eucariotes most of mRNA is monocistronic having an elevated quantity of mRNA-poli A, which is not translated, being broken or chemically modified inside the cells; it is dificult to compare and discuss our results due to lack of published information. The results obtained through these analyses add to the observations made by Paschoal (1991) leading us to think that the alterations in the nucleic acids content in the albumen gland-ovotestis complex of B. similaris infected with sporocysts of E. coelomaticum, are not responsible for snail castration, since there is an inhibition of oviposition, but the gametogenesis is not inhibited. We consider that the castration in this case is partial, and nutritional. ACKNOWLEDGMENTS
To Prof. Edna Maria Gomes, Departamento de CiEncias Fisiologicas, UFRRJ, for providing the laboratory conditions and for suggestions during the biochemical analysis. REFERENCES Amato SB, da Rosa VLM 1982. Observacoes sobre a biologia de Lymnaea (Pseudosuccinea) columella Say. Numero de ovos por massa ovigera relacionado ao tamanho do caramujo. Ci Cul 34: 1640-1641. Baudoin M 1975. Host castration as parasitic strategy. Evolution 29: 335-352.
Brumpt E 1941. Observations biologiques diversees concernat Planorbis (Australorbis) glabratus, hote intermediare de Schistosoma mansoni. Ann Parasitol 18: 9-45. Cheng TC, Sullivan JT, Harris KR 1973. Parasitic castration of the marine prosobranch gastropod Nassarius obsoletus by the sporocyst of Zoogonus rubellus (Trematoda): histopathology. J Invertebr Pathol 21: 183-190. Crews AE, Yoshino TP 1989. Schistosoma mansoni: effect of infection on reproduction and gonadal growth in Biomphalaria glabrata. Exp Parasitol 68: 326-334. Frantz MA, Mossmann RL 1989. Alimentacao de Helix aspersa Muller (Gastropoda-Helicidae) em cativeiro. Acta Biol Leopoldensia 11: 227-233. Loker ES 1979. Effects of Schistosomatium douthitii infection on the growth, survival, and reproduction of Lymnaea catascopium. J Invertebr Pathol 34: 138-144. Mattos Jr DG, Amato SB, Vianna SS da S 1987. Identificacao do segundo hospedeiro intermediario de Eurytrema coelomaticum (Trematoda, Dicrocoeliidae) na regiao do Vale do Paraiba, Sao Paulo. Arq Univ Fed Rur do Rio de J 10: 69-81. McClelland G, Bourns TKR 1969. Effects of Trichobilharzia ocellata on growth, reproduction, and survival of Lymnaea stagnalis. Exp Parasitol 24: 137-146. Pan CT 1965. Studies on the host-parasite relationships between Schistosoma mansoni and the snail Australorbis glabratus. Am J Trop Med Hyg 14: 931-976. Paschoal SV 1991. Eurytrema coelomaticum (Giard et Billet, 1892) Looss, 1907 em Bradybaena similaris - Rota de Migracao e ConsequEncias do Parasitismo sob os Enfoques Biologico e Histologico. Msc Thesis, UFRRJ, Itaguai, RJ. 94pp. Schmidt G, Thannhauser SJ 1945. A method for determination of desoxyribonucleic acid, ribonucleic acid, and phosphoproteins in animal tissues. J Biol Chem 161: 83-89. Sluiters JF 1981. Development of Trichobilharzia ocellata in Lymnaea stagnalis and the effects of infection on the reproductive system of the host. Z Parasitenkd 64: 303-319. Sluiters JF, Brussaard CM, Meuleman EA 1980. The relationship between miracidial dose, production of cercariae, and reproductive activity of the host in the combination Trichobilharzia ocellata and Lymnaea stagnalis. Z Parasitenkd 63: 13-26. Sturrock BM 1966. The influence of infection with Schistosoma mansoni on the growth rate and reproduction of Biomphalaria pfeifferi. Ann Trop Med Parasitol 60: 187-197. Sturrock BM, Sturrock RF 1970. Laboratory studies of the host-parasite relationship of Schistosoma mansoni and Biomphalaria glabrata from St. Lucia, West Indes. Ann Trop Med Parasitol 63: 357-363. Sullivan JT, Cheng TC, Howland H 1985. Studies on parasitic castration of Ilyanassa obsoleta (Mollusca: Gastropoda) by several marine trematodes. Trans Amer Microsc Soc 104: 154-171. Tang CC 1950. Studies on the life history of Eurytrema pancreaticum Janson, 1889. J Parasitol 36: 559-573. van Elk R, Joose J 1981. The UDP-galactose 4-epimerase of the albumen gland of Lymnaea stragnalis and the effects of photoperiod, starvation and trematode infection on its activity. Comp Biochem Physiol [B] 70: 45-52. Vieira S 1986. Introducao a Bioestatistica. Editora Campus, Rio de Janeiro, RJ. 294pp. Villela GG, Bacila M, Tastaldi H 1977. Tecnicas e experimentos de Bioquimica. Editora Guanabara Koogan S.A., Rio de Janeiro, RJ. 552pp. Webe G, James C 1972. Host-parasite relationships of Bulinus globosus and Bulinus truncatus with strains of Schistosoma haematobium. J Helminthol 46: 185-199. Wijdenes J, van Elk R, Joose J 1983. Effects of two gonadotropics hormones on polysacharides synthesis in the albumen gland of Lymnaea stagnalis, studied with the organ culture technique. Gen Comp Endocrinol 51: 263-271. Wilson RA, Denison J 1980. The parasitic castration and gigantism of Lymnaea truncatula infected with larval stages of Fasciola hepatica. Z Parasitenkd 51: 263-271. Wright CA 1966. The pathogenesis of heminths in molluscs. Helminthol Abstr 35: 207-224.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc95129a.gif] |
| |||||||||

{kind=link}