 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Existence of a Sex Pheromone in Triatoma infestans (Hemiptera: Reduviidae): I. Behavioural Evidence Gabriel Manrique/*, Claudio R Lazzari Departamento de Ciencias Biologicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, (1428) Buenos Aires, Argentina
Code Number: OC95131
Size of Files:
Text: 13K
Graphics: Line Drawings (gif) 28K
Assembling behaviour associated with mating was investigated
in Triatoma infestans. The spatial distribution of
both sexes was observed by video films, in the presence or
absence of a copulating pair. Males aggregated around
copulating pairs. Females did not exhibit this behaviour and
their mean spatial density remained unaffected. Spontaneous
aggregation tendency was observed in males in the absence of a
copulating pair, but the temporal course significantly
differed from that observed in the presence of a mating pair.
Results support the existence of an aggregation signal that is
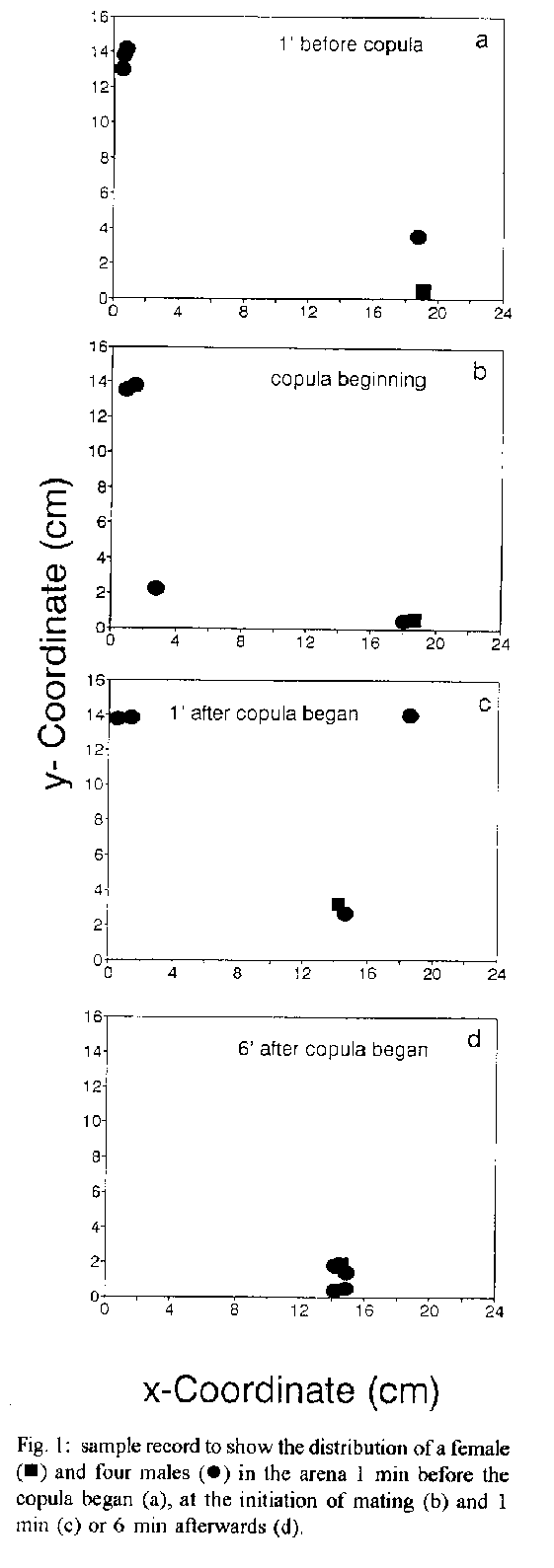
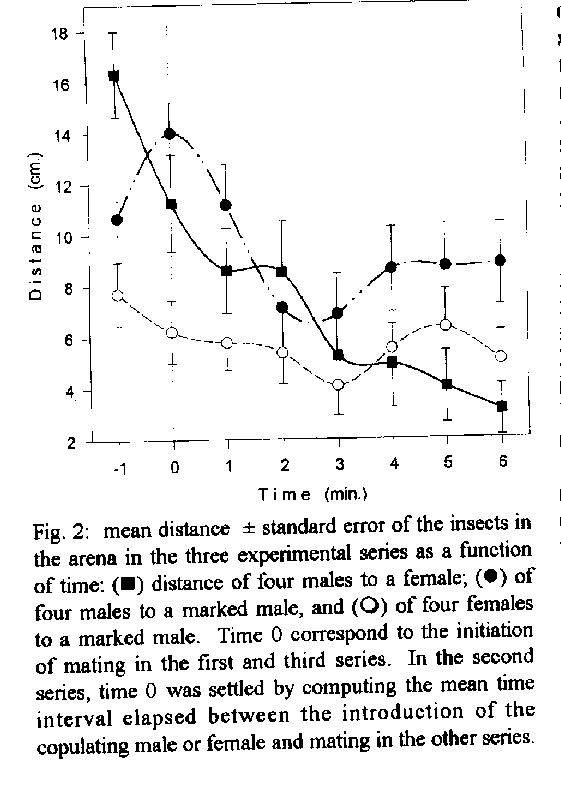
released during mating, affecting the behaviour of males.Key words: Triatominae - pheromones - communication - aggregation - Chagas' disease The haematophagous bug, Triatoma infestans, is the principal vector of Trypanosoma cruzi parasites, the causative agent of Chagas' disease in South America. Understanding chemical signals involved in the intraspecific communication of this species and other Triatominae may provide mechanisms for the development of control strategies. Intraspecific communication has been demonstrated in T. infestans in relation to assembling behaviour (Schofield & Patterson 1977, Lorenzo Figueiras et al. 1994). The existence of pheromones affecting sexual behaviour in Triatominae bugs has been investigated. Evidence about chemical attraction between males and females was reported by Neves and Paulini (1981) for T. infestans and Panstrongylus megistus, and by Antich (1968) for Rhodnius prolixus. In addition, the existence of an assembling factor released during the copula affecting the spatial distribution of males, was investigated in triatomines. Up to now, its occurrence was reported in R. prolixus by Baldwin et al. (1971) who stated that a sex pheromone is released during the copula, promoting the aggregation of males around the mating pair. On the other hand, the apparent absence of pheromones in T. infestans affecting sexual behaviour was reported by Schofield and Moreman (1976). Contradictory results were reported for Triatoma mazzottii (Ondarza et al. 1986, 1987, Rojas et al. 1991). In the present study we address the question about the existence of a chemical stimuli in T. infestans which is released during copula, inducing males to assemble similarly to R. prolixus. The modification of the spatial distribution of individuals of both sexes in the presence or absence of a copulating pair, was evaluated. MATERIALS AND METHODS The study was designed to quantify the aggregation of males and females, in the presence of a mating pair. Virgin adults with a high probability of copulating (Manrique & Lazzari 1994) were used. Insects were reared at 28 C and fed on heparinized goat blood, using an artificial feeder (Nunez & Lazzari 1990). Experiments were conducted in an experimental arena, consisting of a rectangular glass recipient (24cm x 16cm x 6cm high). A piece of filter paper (24cm x 16cm) was used as substratum. Temperatures were maintained at 25+/-1 C and diffuse illumination of 35 lux intensity was provided by a tungsten lamp. Three experimental series were conducted. Individuals were released in the centre of the arena in all series. In the first one, four males were released in the experimental arena and after 15 min, a female was introduced (4 replicates). In the second one, four males were released in the experimental arena and after 15 min, a marked male was introduced (5 replicates). In the third one, four females were released in the experimental arena and after 15 min, a male was introduced (4 replicates). The behaviour of the insects was recorded by means of video films. The x- y- coordinates corresponding to each insect (centre of pronotum as reference) were read frame by frame and fed to a computer with the aid of a digitizing device. The distance between each insect in the arena and the individual introduced afterwards was computed for all experimental series. A quantitative analysis of the displacements of these insects was performed by studying the variations in distance to the reference individual as a function of time, before, during and after the copula. RESULTS While the copula took place, the males in the arena exhibited a tendency to aggregate close to the mating pair (Fig.1). The distance between the males and the mating female decreased over time and diminished progressively after the copula began (Fig. 2) (series 1). Significant differences in the distribution of the insects between series (F= 15.2, P<0.001) as well as along time (F= 6.7, P<0.001) were observed (Two way ANOVA). A significant interaction between factors was also observed (F= 2, P=0.013), demonstrating a differential variation of the measured parameter over time in all three experimental groups. When the aggregation of males on a marked male was tested (series 2), the distance to the marked one also diminished along time, demonstrating a spontaneous tendency to aggregate in males, even in the absence of a mating pair (Fig. 2). However, there is a significant difference in the dynamics of the behaviour when compared in both situations, i.e. in the presence and in the absence of a mating pair (Scheffe pairwise comparison, P= 0.02). Moreover, the distance between the males and the introduced female decreased at a faster rate when compared with the decrease in distance from males to the introduced marked male. In the presence of a mating pair, the aggregation had a directional component, with males assembling around the mating pair, whereas in the absence of mates, aggregation was random.
When the aggregation of females on a mating couple was tested (series 3), no significant change in the distance to the copulating male was observed, i.e females did not aggregate (Fig. 2). The dynamics of the distribution significantly differed from the other two situations (Scheffe test, P<0.05 in both cases). DISCUSSION Our results support the existence of an aggregation stimuli produced during mating, resulting in neighbouring males, but not females, aggregating around a mating pair. Superimposed to the apparent spontaneous tendency of males to randomly assemble, detailed analysis demonstrated the effect of a sex signal during copula of a mating pair. Males tend to assemble in the absence of any female, but the dynamics of their aggregation behaviour was different in both situations, i.e. in the presence or in absence of a mating pair. In the first case, the malesÆ behaviour would be mediated by a spontaneous tendency to maintain a high body contact with the substratum (thigmotaxis) (Lorenzo Figueiras et al. 1994) , whereas in the second case chemical cues would be involved (de Brito Sánchez et al. 1995), although thigmotaxis can not be excluded. Previous reports concerning sexual behaviour of T. infestans suggested the absence of male aggregation sex pheromones (Schofield & Moreman 1976). Different experimental approaches could ascribe for the different results. Whereas Schofield and Moreman (1976) photographed the insects every 5 min for 18 hr, our video films provided a continuous record. In some of our assays the mating lasted for about 6 min or less, a period of time near to the sample interval chosen by those authors. This behavioural study along with the electrophysiological evidence brought by de Brito Sánchez et al. (1995) strongly supports the existence of sex pheromones released by one or both members of a copulating pair, which in turn modify the behaviour of other males. The biological significance of the sex signal of T. infestans and the modification of males' behaviour is far from being understood. Some authors (Baldwin et al. 1971, Schofield 1979, Ondarza et al. 1986, 1987) ascribed to this signal in other Triatominae as an aphrodisiac function. This interpretation is based on: (1) the signal is released during the copula (Ondarza et al. 1986, 1987), and (2) in the presence of a pair, R. prolixus males attempted to copulate with other males (Baldwin et al. 1971). In the first case, it is difficult to understand the significance of behavioural modification for males as a result of mating individuals. If the intention was to “excite” other males, to induce them to copulate with any available female, it is not easy to comprehend how it evolved. Concerning the second point, we also observed males that attempted to copulate with other males in the presence of a mating pair. However, this behaviour is observed also in the absence of mating pairs. Some preliminary experiments show that the assembled males copulate one after the other with the same female (Lazzari, unpubl.). Thus, the sex signal could be related to polyandry, a quite rare and puzzling mating system in insects (Thornhill & Alcock 1983). The analysis of the temporal course of the aggregation of males around a mating couple constitute a simple and specific bioassay, useful for testing the different fractionating stages of the putative pheromone, e.g. sex, glands, chemical fractions, etc. The adequate isolation and characterisation of the active compound/s may provide mechanisms for the development of control strategies for ChagasÆ disease vectors. ACKNOWLEDGEMENTS To W Farina, G Flores, M Giurfa, P Guerenstein, J Ianowski, T Insausti, R Josens, M Lorenzo, A Lorenzo Figueiras, L Moffatt, J Nunez, F Roces and P Schilman for many helpful discussions and critical reading of the manuscript. REFERENCES Antich AV 1968. Atracion por olor en ninfas e adultos de Rhodnius prolixus. Rev Inst Med Trop y Parasitol 10: 242-246. Baldwin WF, Knight AG, Lynn KR 1971. A sex pheromone in the insect Rhodnius prolixus (Hemiptera: Reduviidae). Can Entomol 103: 18-22. de Brito Sanchez MG, Manrique G, Lazzari CR 1995. Existence of a sex pheromone in Triatoma infestans (Hemiptera: Reduviidae): II. Electrophysiological correlates. Mem Inst Oswaldo Cruz 90: 649-651. Lorenzo Figueiras AN, Kenigsten A, Lazzari CR 1994. Aggregation in the haematophagous bug Triatoma infestans: chemical signals and temporal pattern. J Insect Physiol 40: 311-316. Manrique G, Lazzari CR 1994. Sexual behaviour and stridulation during mating in Triatoma infestans (Hemiptera: Reduvidae). Mem Inst Oswaldo Cruz 89: 629-633. Neves DP, Paulini E 1981. Atracao sexual em Panstrongylus megistus e Triatoma infestans (Hemiptera, Reduviidae) por feromonio. Rev bras Ent 25: 301-306. Nunez JA, Lazzari CR 1990. Rearing of Triatoma infestans Klug (Het., Reduviidae) in the absence of a live host. I. Some factors affecting the artificial feeding. J Appl Entomol 109: 87-92. Ondarza RN, Gutierrez-Martinez A, Malo EA 1986. Evidence for the presence of sex and aggregation pheromones from Triatoma mazzottii (Hemiptera: Reduviidae). J Econom Entomol 79: 688-692. Ondarza RN, Gutierrez-Martinez A, Malo EA, Rojas JC 1987. Actividad afrodisiaca de la feromona sexual de Triatoma mazzottii Usinger. Southwest Entomol 12: 327-333. Rojas JC, Ramirez-Rovelo A, Cruz-Lopez L 1991. Search for a sex pheromone in Triatoma mazzottii (Hemiptera: Reduviidae). J Med Entomol 28: 469-470. Schofield CJ 1979. The behaviour of Triatominae (Hemiptera: Reduviidae): A review. Bull Entomol Res 69: 363-379. Schofield CJ, Moreman K 1976. Apparent absence of a sex attractant in adult Triatoma infestans (Klug), vector of Chagas' Disease. Trans R Soc Trop Med Hyg 70: 165-166. Schofield CJ, Patterson JW 1977. Assembly pheromone of Triatoma infestans and Rhodnius prolixus nymphs (Hemiptera: Reduviidae). J Med Entomol 13: 727-734. Thornhill R, Alcock J 1983. The evolution of insect mating systems. Harvard University Press - Cambridge, USA. 547 pp.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc95131b.gif] [oc95131a.gif] |
| |||||||||

{kind=link}
{kind=link}