 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Existence of a Sex Pheromone in Triatoma infestans (Hemiptera: Reduviidae): II. Electrophysiological Correlates Maria G de Brito Sanchez, Gabriel Manrique*, Claudio R Lazzari Departamento de Ciencias Biologicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, (1428) Buenos Aires, Argentina
Code Number: OC95132
Size of Files:
Text: 12K
Graphics: Line Drawings (gif) 17K
The stimulus provided by a copulating pair of Triatoma
infestans significantly affects the electrical activity of
the nervous system of Triatoma infestans.
Electrophysiological recordings were performed on stationary
adult males presented with stimuli of an air current carrying
odors from males, females, non-copulating pairs and mating
pairs. The electrophysiological response was characterized by
the low frequency occurrence of biphasic compound impulses. A
significant increase in the frequency of the impulses occurred
in stationary males when exposed to air currents of mating
pairs, when compared to that evoked by a clean air stream.
Analysis of the time course of the assays, showed that the
electrophysiological activity during the copula was higher
than prior to or after copula. The electrophysiological
evidence presented here strongly supports the existence of
pheromone(s) released by one or both sexes during mating and
which is perceived by male chemoreceptors located on the
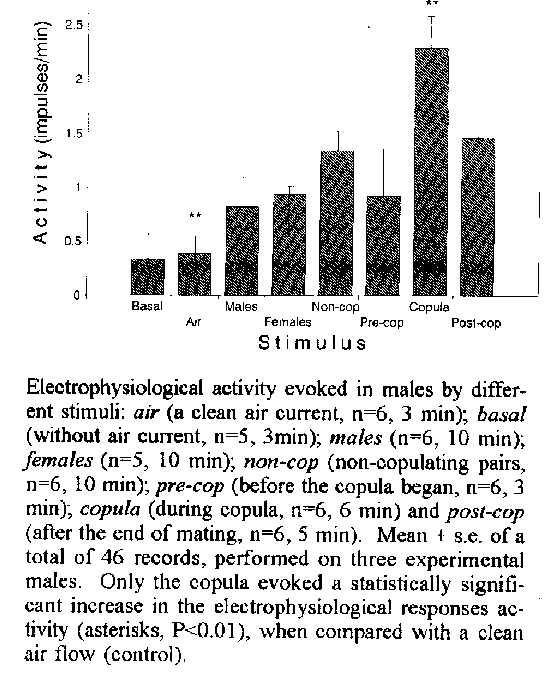
antennae.Key words: Triatominae - pheromones - communication - electrophysiology - Chagas' disease Triatoma infestans (Klug) is of medical importance, since it is a vector for the transmission of the flagellate Trypanosoma cruzi, the causative agent for american trypanosomyasis. In spite of this, relatively little is known about the chemical signals involved in its intraspecific communication. These chemosensory responses are crucial for developing alternative control measurements for this species (e.g., Schofield & Patterson 1977, Nunez 1987, Cruz Lopez et al. 1993, Lorenzo Figueiras et al. 1994). The existence of pheromones affecting sexual behaviour in Triatominae bugs, has been reported for Rhodnius prolixus (Baldwin et al. 1971). Whereas the apparent absence of sex pheromones has been reported for Triatoma infestans (Schofield & Moreman 1976), contradictory evidence has been reported for Triatoma mazzottii (Ondarza et al. 1986, 1987, Rojas et al. 1991). For R. prolixus, this pheromone is released during the copula and promotes the assembling of males around the mating couple. For T. infestans, preliminary observations, based on behavioural evidence, reported that males aggregate around a copulating pair and the existence of a sex pheromone released by one or both members of the pair was suggested (Manrique & Lazzari 1995). This study investigated the events associated with the behavioural response of T. infestans males based on electrophysiological signals of antennal chemoreceptors. We addressed the question of whether the stimulus provided by a copulating pair significantly affects the electrical activity in the proximity of the antennal nerve. Materials and Methods Sexually mature T. infestans, reared at 28 C and fed weekly on heparinized goat blood using an artificial feeder were used (Nunez & Lazzari 1990). Since only males evince a behavioural response associated with a copulating pair (Manrique & Lazzari 1995), electrophysiological records were conducted in this sex. Insects were ventrally fastened with plasticine to a Petri dish with their antennae remaining free. A small window was opened on the antenniferous tubercle under a dissection microscope and the cuticle was removed, exposing the antennal nerve before its branches (Insausti 1994). An indifferent tungsten electrode was inserted in the compound eye. The recording tungsten electrode was carefully placed on the antennal nerve and verified under the dissection microscope. The response obtained was filtered, amplified and displayed on a storage oscilloscope from which it was videorecorded for posterior analysis. This method was chosen because of the kind of impulses recorded, that allowed them to be recorded in videotapes without loss of information (see below). Concurrently, behavioural responses observed in the copulating pairs used as stimuli were recorded so that the occurrence of the copula could be correlated with electrophysiological events. Constant air stream (10ml/min), produced by a pump and conducted through a plastic box (10cm x 10cm x 6cm high) where the stimuli were placed, flowed over the antennae of the fixed male through rubber tubing (5mm diam., 40cm long from the pump to the box, and 50cm from the box to the insect). The air flow was regulated through a system of valves and verified by means of a flowmeter. The basal activity was measured in the absence of any stimulus (i.e. without airstream) and also in the presence of a clean air stream (no insects in the box). Pairs of adults, prone to copulate (Manrique & Lazzari 1994), were placed in the box through which the air stream passed to the experimental males from which recordings were obtained. The following stimuli were presented inside the box: (a) two males, (b) two females or, (c) a male and a female. The latter were monitored by direct observation throughout the experiment, to determine whether copulation took place. When mating occurred, the records were sorted in intervals corresponding to the time prior to (3 min), during and after (5 min), copula. The frequency of the biphasic compound impulses was computed for different experimental conditions and compared with the activity observed in males in the presence of a clean air current (control) by means of a paired samples t-test. RESULTS The electrophysiological response was characterized by the low frequency occurrence (1-12 per min) of biphasic compound impulses (0.4 mV peak to peak). No significant variation in activity was observed for males without stimulus (basal activity vs. clean air stream). Similar results were obtained when fixed adults males were presented with a clean air of constant flow conducted through the container with two males, two females or a non-copulating pair (Fig.). Nevertheless, a significant increase in the frequency of impulses occurred whenever a pair copulated in the box (t = 10.15, P<0.01). Analysis over time for the assays, showed that the activity during the copula was higher than in the minutes prior to and after copula (Fig.). DISCUSSION The possible existence of a chemical sex signal in T. infestans, employing an electrophysiological approach, i.e. by searching for changes at the nervous level that correlate with sexual behaviour was investigated. Significant variations in electrical activity associated exclusively with copulation were found in males of this species. Other stimuli tested (i.e., males, females or non-copulating pairs) failed to evoke such electrophysiological responses. The recording modality did not allow us to distinguish between observed activity originating in sensory inputs or evoked motor outputs. However, this fact appears to be not relevant in the present context, since sensory as well as motor activity (e.g. antennal movements) occur in the presence of attractive stimuli in this species ( Lazzari & Nunez 1989, Flores & Lazzari 1993). The electrophysiological evidence presented here strongly supports the existence of pheromone(s) released by one or both sexes during mating since the olfactory modality is the only that can explain the observed changes in the electrophysiological responses recorded. Moreover, these results are correlated with the aggregation behaviour performed by males around a couple, that takes place in the presence of a mating couple (Manrique & Lazzari 1995). According to Manrique and Lazzari (1995) these behavioural responses only occur in males and not in females. Since studies were conducted only in males, it is not possible, to discern differences between sexes, i.e., sensilla, receptors, central filters or behavioural outputs. Both, behavioural and electrophysiological approaches provides relevant information about the putative chemical signal and its biological role. The characterization of the pheromone(s) involved would be useful for the developing control strategies of ChagasÆ disease vectors. ACKNOWLEDGEMENTS To W Farina, G Flores, M Giurfa, P Guerenstein, J Ianowski, T Insausti, R Josens, M Lorenzo, A Lorenzo Figueiras, L Moffatt, J Nunez F Roces and P Schilman for many helpful discussions and critical reading of the manuscript. To D Varju for kindly donating the electrophysiological setup. REFERENCES Baldwin WF, Knight AG, Lynn KR 1971. A sex pheromone in the insect Rhodnius prolixus (Hemiptera: Reduviidae). Can Entomol 103: 18-22. Cruz Lopez L, Malo EA, Rojas JC 1993. Aggregation pheromone in five species of Triatominae (Hemiptera: Reduviidae). Mem Inst Oswaldo Cruz 28: 535-539. Flores GB, Lazzari CR 1993. Bilateral integration of antennal inputs during thermal orientation in Triatoma infestans. Mem Inst Oswaldo Cruz 88 Suppl.: 250. Insausti TC 1994. Nervous system of Triatoma infestans. J Morphol 221: 343-359. Lazzari CR, Nunez JA 1989. The response to radiant heat and the estimation of the temperature of distant sources in Triatoma infestans. J Insect Physiol 35: 525- 529. Lorenzo Figueiras AN, Kenigsten A, Lazzari CR 1994. Aggregation in the haematophagous bug Triatoma infestans: chemical signals and temporal pattern. J Insect Physiol 40: 311-316. Manrique G, Lazzari CR 1995. Existence of a sex pheromone in Triatoma infestans (Hemiptera: Reduviidae): I. Behavioural evidence. Mem Inst Oswaldo Cruz 90: 645-648.. Manrique G, Lazzari CR 1994. Sexual behaviour and stridulation during mating in Triatoma infestans (Hemiptera: Reduviidae). Mem Inst Oswaldo Cruz 89: 629-633. Nunez JA 1987. Behaviour of Triatominae bugs, p.1-28. In ChagasÆ Disease Vectors, Vol II, RR Brenner, AM Stoka (eds). CRC Press, Florida. Nunez JA, Lazzari CR 1990. Rearing of Triatoma infestans Klug (Het., Reduviidae) in the absence of a live host. I. Some factors affecting the artificial feeding. J Appl Entomol 109: 87-92. Ondarza RN, Gutierrez-Martinez A, Malo EA 1986. Evidence for the presence of sex and aggregation pheromones from Triatoma mazzottii (Hemiptera: Reduviidae). J Econom Entomol 79: 688-692. Ondarza RN, Gutierrez-Martinez A, Malo EA, Rojas JC 1987. Actividad afrodisiaca de la feromona sexual de Triatoma mazzottii Usinger. Southwest Entomol 12: 327-333. Rojas JC, Ramirez-Rovelo A, Cruz-Lopez L 1991. Search for a sex pheromone in Triatoma mazzottii (Hemiptera: Reduviidae). J Med Entomol 28: 469-470. Schofield CJ, Moreman K 1976. Apparent absence of a sex attractant in adult Triatoma infestans (Klug), vector of ChagasÆ Disease. Trans R Soc Trop Med Hyg 70: 165-166. Schofield CJ, Patterson JW 1977. Assembly pheromone of Triatoma infestans and Rhodnius prolixus nymphs (Hemiptera: Reduviidae). J Med Entomol 13: 727-734.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc95132a.gif] |
| |||||||||

{kind=link}