 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biologia do Triatoma nitida Usinger, 1939 em Laboratorio (Hemiptera: Reduviidae) Cleber Galvao, Jose Jurberg, Vanda Cunha, Rubens Pinto de Mello* Laboratorio Nacional e Internacional de Referencia em Taxonomia de Triatomineos *Laboratorio de Diptera, Departamento de Entomologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
Code Number: OC95134
Size of Files:
Text: 28K
Graphics: Line Drawings (gif) 51K
Biology of Triatoma nitida Usinger, 1939 under
Laboratory Conditions -Triatoma nitida is a wild
species occurring in Mexico and Central America. In order to
establish the length of its life cycle and transmission
potential, the following parameters were observed: the
incubation period, the interval between hatching, or moulting,
and the first feeding; the number of blood meals and the time
of development. The time-lapse before the bite, the length of
feeding and the interval between the end of the blood meal and
defecation, as well as the site of defecation were also
analyzed. Average length of the egg incubation period was 18.2
days. Time interval between the food source offering and the
bite was less than 4 min in 60% of the analyzed cases, except
in the fifth instar, where only 38% of the insects began
feeding in less than 5 min. The blood-sucking period was long
and rising until the fifth instar , decreasing in adults, and
ranging from 1 min to 2 and a half hours. Only 26% of the
blood meals were followed by defecations within 20 min. The
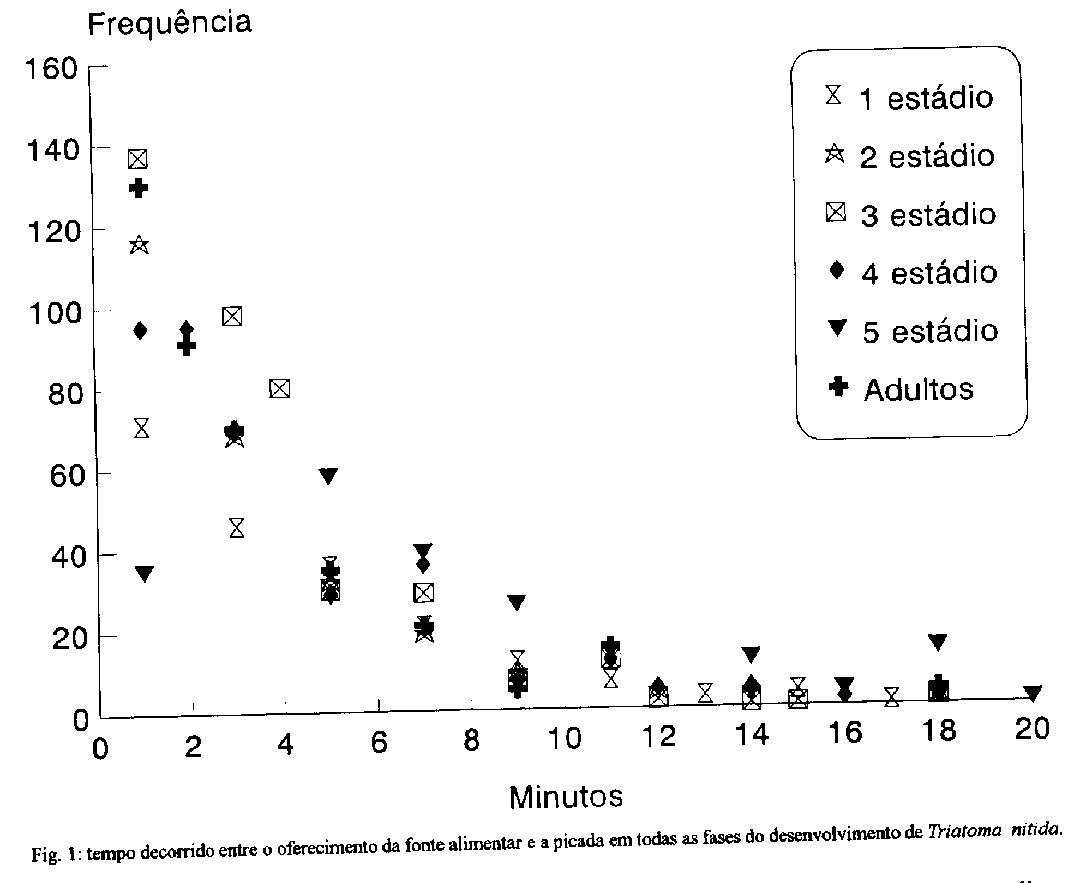
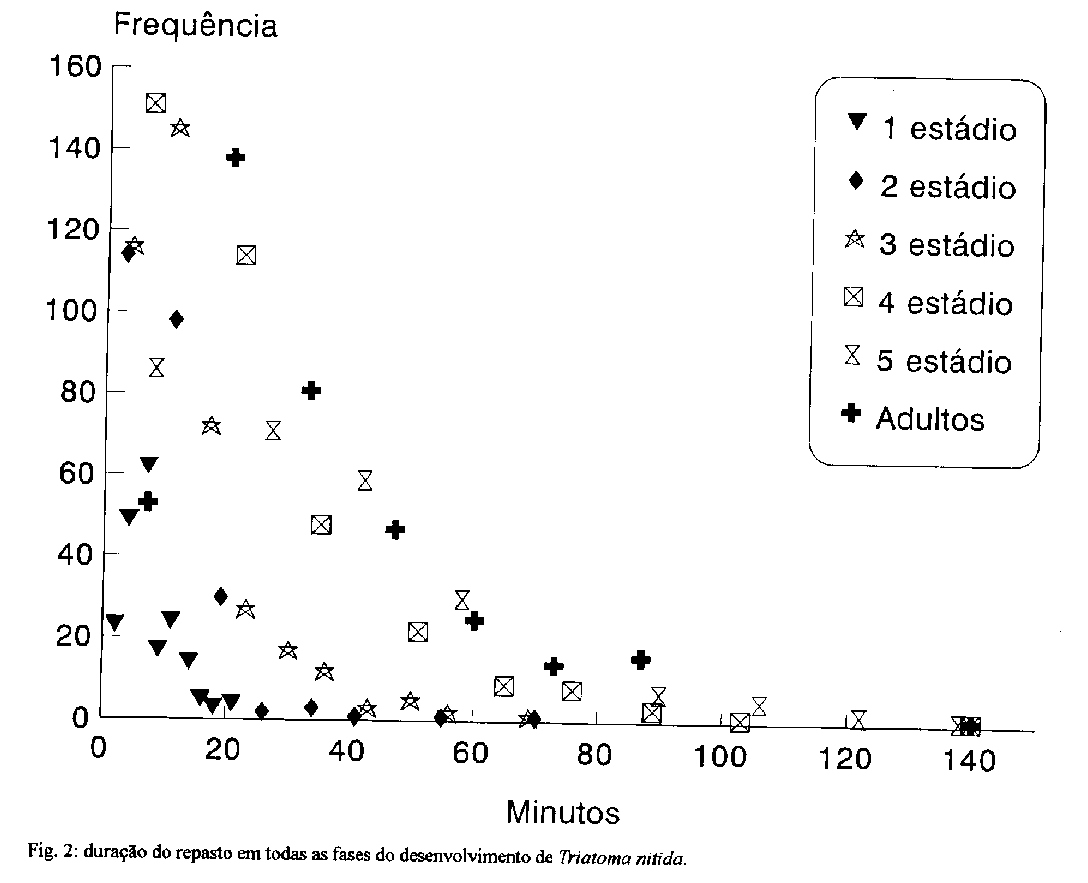
average length of the life cycle was 897.5 days.Key words: life cycle - Triatoma nitida - Triatominae - Chagas' disease Triatoma nitida Usinger, 1939 e uma especie silvestre, de ocorrEncia assinalada na Costa Rica, Guatemala, Honduras e Mexico. Dias (1952) publicou uma lista da distribuicao das especies centro-americanas e assinalou as localidades onde foi verificada a infeccao de triatomineos pelo Trypanosoma cruzi. T. nitida foi encontrado infectado pelo flagelado na Guatemala, porem em Honduras nenhum dos especimes examinados apresentou infeccao. Leon (1959) chamou atencao para o alto grau de infeccao dessa especie na Guatemala, onde 83% dos especimes capturados encontravam-se positivos para T. cruzi. Gonzalez-Angulo e Ryckman (1967) encontraram, pela primeira vez no Mexico (Yucatan), representantes dessa especie. Sob o ponto de vista morfologico e taxonomico, esse triatomineo foi analisado nos trabalhos de Lent e Wygodzinsky (1979), Jurberg et al. (1991), Lent e Jurberg (1992), entretanto sua biologia, hospedeiros naturais e habitats sao pouco conhecidos. Com intuito de ampliar os parametros que permitem a definicao precisa dessa especie, registrou-se o periodo de desenvolvimento e avaliou-se o potencial vetorial em condicoes de laboratorio. Dentro deste proposito, observaram-se: o periodo de incubacao, o intervalo entre a eclosao ou muda e o primeiro repasto; o numero de repastos e o periodo de desenvolvimento de ovo a adulto em 92 especimes. A observacao de 1856 repastos sanguineos possibilitou o registro do intervalo entre o oferecimento da fonte alimentar e a picada; o tempo de duracao do repasto; o intervalo decorrido entre o fim do repasto e a defecacao; e o local da defecacao. MATERIAIS E METODOS Os insetos estudados eram provenientes de uma colonia aclimatada no insetário do Laboratorio Nacional e Internacional de ReferEncia em Taxonomia de Triatomineos, do Departamento de Entomologia do IOC, onde sao mantidos em temperatura ambiente com alimentacao quinzenal em pombos (Columba livia). Quinze especimes de 5^o estadio foram retirados da colonia e mantidos em um cristalizador de vidro transparente, de 20 cm de altura por 17 cm de diametro, e observados diariamente ate a ocorrEncia da muda imaginal. Destes, cinco femeas e tres machos foram transferidos para um recipiente metalico de 21 cm de altura por 16 cm de diametro, com o fundo de tela de arame, visando facilitar a coleta dos ovos. Os ovos coletados diariamente, eram agrupados de acordo com a data da postura e aclimatados em estufa B.O.D. a temperatura de 28 +/- 1 C e 80 +/- 5% de U.R., com fotoperiodo controlado, alternando-se 12 h de luz com 12 de obscuridade. Dos 135 ovos separados, 92 eclodiram. As ninfas recem emergidas foram individualizadas em frascos de Borrel (9 cm de altura x 3 cm de diametro) numerados, forrados no fundo por papel de filtro e com uma tira do mesmo papel para absorver o excesso de umidade e proporcionar maior area de deslocamento aos insetos. Os frascos foram fechados com telas de náilon fixadas por elasticos. A alimentacao foi oferecida diariamente ate a ocorrencia do primeiro repasto de cada fase, passando entao a ser semanal. Foi realizada em camundongos albinos (Mus musculus), que tinham as patas imobilizadas por meio de fita adesiva. No dorso do animal afixava-se uma tira de papel para servir de apoio ao inseto durante o repasto. O tempo de oferecimento da fonte alimentar foi de 20 min., sendo o tempo de succao rigorosamente cronometrado, desde o momento da picada; apos o termino do repasto, aguardavam-se 20 min. para verificar o local (sobre a fonte ou nao) e o tempo decorrido ate o momento da defecacao. RESULTADOS E DISCUSSAO Ovos - O periodo de incubacao dos 92 ovos observados variou de 13 a 21 dias, com media de 18.2 (S = 1.49 e S2 = 2.24). Resultados semelhantes foram obtidos por Carcavallo e Martinez (1972) para T. delpontei Romana & Abalos, 1947; por Rabinovich (1972) para T. infestans (Klug, 1834) e por Silva e Silva (1991) para T. dimidiata (Latreille, 1811) criados em condicoes similares. Realizacao do primeiro repasto - Atraves do oferecimento diario da fonte alimentar foi possivel registrar o intervalo decorrido entre a eclosao, ou muda, e o primeiro repasto. Os resultados obtidos mostraram que o intervalo foi crescente, apesar de a media do 2^o estadio ter sido menor que a do 1o. Todas as ninfas precisaram de ao menos um dia de 'repouso' pre-alimentar apos a eclosao, ou muda, e os adultos de pelo menos dois dias (Tabela I). Lafeta (1966) demonstrou que, a temperatura de 25 C e 80% de U.R., as ninfas de 1^o estadio de Panstrongylus megistus (Burmeister, 1835) so se alimentaram a partir do quarto dia de vida, e que no sexto dia a alimentacao foi realizada por 80% dos especimes. Segundo Perlowagora-Szumlewicz (1969), a primeira alimentacao do T. infestans teria importancia vital para o surgimento precoce dos adultos, e que o quarto dia apos a eclosao seria o ideal para promover esta possibilidade. Leopoldo-e-Silva et al. (1990) testaram esta hipotese em ninfas de T. infestans e Rhodnius prolixus Stal, 1859 e demonstraram que nao houve diferenca significativa nos resultados obtidos para os insetos alimentados no 4o, no 8o e no 12o dia. Tempo decorrido entre o oferecimento da fonte alimentar e a picada - O registro do tempo decorrido entre o oferecimento da fonte e a picada permitiu avaliar a agressividade e voracidade na procura pelo alimento. T. nitida demonstrou pouca agressividade, apesar disso, em cerca de 60% dos casos observados, o intervalo entre o oferecimento e a picada foi inferior a 4,5 min.; e em apenas 7% este intervalo foi superior a 10 min. Este foi o padrao para todas as fases, exceto para o 5o estádio, onde apenas 38% aceitaram a alimentacao em menos de 5 min. e 16% ultrapassaram os 10 min. ( Fig. 1). Deve-se considerar que a variabilidade no periodo de tempo que antecedeu a picada, certamente sofreu a influEncia do manuseio e da mudanca de temperatura e umidade ocasionadas pela retirada dos insetos da estufa para possibilitar as observacoes. Tempo de succao e eliminacao das dejecoes - A realizacao do repasto sanguineo possibilita o contato entre o vetor e o hospedeiro; a consequente defecacao e o meio mais comum de exposicao do hospedeiro ao T. cruzi. Estudos sobre o tempo de alimentacao e tempo de defecacao sao de grande importancia para ampliar os conhecimentos sobre a transmissao do T. cruzi e apontar as especies mais capacitadas a desempenhar com eficiencia seu potencial vetorial. Je em 1949 Dias e Chandler alertavam para a importancia dos habitos de defecacao dos triatomineos, porem, foi Wood (1951) o primeiro a desenvolver experimentos sobre tais habitos. A duracao do repasto foi crescente do 1^o ate o 5^o estadio; na fase adulta sofreu pequena reducao e ficou mais proxima dos resultados obtidos para o 4o estadio (Fig. 2). Esta reducao no tempo de succao dos adultos esta diretamente relacionada a quantidade de sangue ingerida. Esse aspecto ja foi demonstrado por Pippin (1970), em T. sanguisuga Usinger, 1944, T. gerstaeckeri (Stal, 1859) e R. prolixus; por Wood (1976), em T. protracta (Uhler, 1894); por Zeledon et al. (1977) em R. prolixus, T. infestans e T. dimidiata; por Zarate et al. (1984) para T. barberi Usinger,1939 e mais recentemente por Rocha et al. (1994) para R. pictipes Stal, 1872. TABLE 1. Intervalo de tempo (em dias) decorrido entre a enclosao, ou muda de Triatoma nitida, e o primeiro repasto
--------------------------------------------------- Fases Min. Max. X S S^2 N --------------------------------------------------- 1^o estadio 1 22 5,6 3,36 11,34 74 2^o estadio 1 13 3,7 2,45 6 72 3^o estadio 1 29 5,7 4,46 19,92 68 4^o estadio 1 26 8,3 5,56 30,97 60 5^o estadio 1 50 10,8 9,08 82,58 56 Adultos 2 55 14,1 10,1 102,17 39 ---------------------------------------------------
TABELA II. Numero de repastos realizados em cada fase do desenvolvimento de Triatoma nitida
--------------------------------------------------- Fases Min. Max. X S N --------------------------------------------------- 1^o estadio 1 5 2,7 0,84 73 2^o estadio 1 8 3,4 1,16 71 3^o estadio 3 12 6,1 1,91 60 4^o estadio 3 18 6,1 2,59 57 5^o estadio 3 14 5,2 2,47 40 Adultos 1 26 10,1 6,65 37 ---------------------------------------------------TABELA III. Duracao das fases de desenvolvimento (em dias) de Triaoma nitida --------------------------------------------------- Fases Min. Max. X S N --------------------------------------------------- 1^o estadio 14 105 54,6 20,32 73 2^o estadio 13 202 60,6 33,44 71 3^o estadio 49 358 180,3 71,8 60 4^o estadio 89 446 228 93,41 57 5^o estadio 60 399 217,1 90,29 40 Adultos 26 457 193,2 115,15 39 ---------------------------------------------------
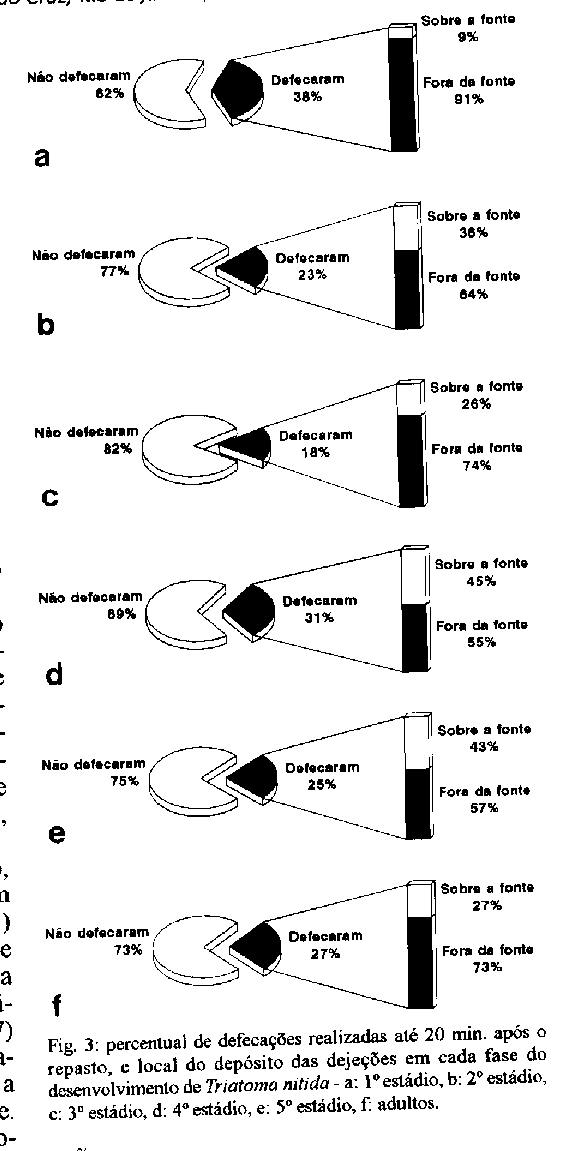
Fig. 3: percentual de defacocoes realizadas ate 20 min. apos o repasto, e local do deposito das dejecoes em cada fase do desenvolvimento de Triatoma nitida- a: 1^o estadio, b: 2^o estadio, c: 3^o estadio, d: 4^o estadio, e: 5^o estadio, f: adultos Fig. 4: percentual de mortalidade das ninfas de Triatoma nitida
O tempo de defecacao e crucial na transmissao do T. cruzi, sendo mais eficazes as especies que defecam durante ou logo apos o repasto. Esse aspecto foi discutido, entre outros, por Dias (1956), Zeledon et al. (1977), Ozete (1981), Zarate et al. (1984), Kirk e Schofield (1987), Costa e Jurberg (1990) e Rocha et al. (1994). Neste experimento, dos 1856 repastos realizados, 484 (26%) foram seguidos de defecacoes no prazo de 20 min., e destas, apenas 152 (31.4%) foram depositadas diretamente sobre a fonte. Os resultados obtidos para cada fase do ciclo encontram-se na Fig.3 e indicam que, sob este aspecto, T. nitida apresenta baixo potencial vetorial. Numero de repastos realizados em cada fase do ciclo de vida - O oferecimento semanal da alimentacao e a manutencao dos especimes que se recusavam a sugar resultaram em grandes variacoes individuais no numero de repastos realizados, porem, deste modo, as caracteristicas individuais foram mantidas, evitando-se a manipulacao dos resultados. A Tabela II mostra os resultados obtidos para este parametro em cada fase do desenvolvimento. Segundo Juarez (1970), a avaliacao do numero de repastos tem grande importancia epidemiologica , uma vez que quanto mais contatos ocorrerem entre vetor e hospedeiro, maior sera a possibilidade de infeccao ou transmissao do T. cruzi. Em T. nitida, o grande numero de repastos realizados aumentaria a probabilidade de infeccao e/ou transmissao, porem na natureza aumentaria tambem a exposicao dos insetos a riscos. Embora algumas especies possam atingir ate o 4o estadio com apenas um repasto por fase, ha que se considerar a influencia de fatores como temperatura e fonte alimentar, alem do stress causado pela manipulacao. Freitas et al. (1967) demonstraram que com alimentacao semanal o ciclo de vida do R. neglectus Lent, 1954 foi mais curto quando comparado com a alimentacao quinzenal. Mc Ewen e Lehane (1993) verificaram que ninfas de 5o estadio de T. infestans alimentadas com mais frequencia tinham seu desenvolvimento acelerado. A quantidade e a fonte de sangue podem alterar, significativamente, alem do tempo de desenvolvimento, a fecundidade e a capacidade de dispersao dos adultos pelo voo (Ekkens 1981, Lehane e Schofield 1981,1982). Costa et al. (1986, 1987), Diotaiuti e Dias (1987) e Costa e Jurberg (1990) demonstraram o melhor desenvolvimento de diversas especies quando alimentadas em camundongos do que em pombos. Duracao do ciclo de vida - T. nitida apresentou um dos mais longos periodos de desenvolvimento ja relatados na literatura. A media de duracao do ciclo, da eclosao a fase adulta (ate a data da morte) foi de 897,6 dias com um periodo minimo de 523 e maximo de 1394 dias. Estes resultados (Tabela III) aproximam-se dos obtidos por Dias (1955) e Souza et al. (1978) para T. sordida (Stal, 1859) e Heitzmann-Fontenelle (1983) para T. brasiliensis. Lent e Jurberg (1969) observando o ciclo do Panstrongylus geniculatus (Latreille, 1811), obtiveram resultados dilatados devido a necessidade de alta umidade para a criacao dessa especie.
A principal vantagem da criacao individual e permitir a observacao detalhada de inumeros parametros biologicos importantes para a determinacao do potencial vetorial da especie. Entretanto, deve-se considerar a possibilidade de efeitos negativos, causados pelo manuseio excessivo e confinamento dos insetos. Tolezano et al. (1984) atribuiram ao excesso de manuseio o aumento da mortalidade e a diminuicao da fertilidade do T. infestans e Costa e Jurberg (1990) recomendaram cautela na aceitacao dos resultados obtidos em criacoes individuais de C. lenti, devido ao stress causado pela manuseio, ja que esta especie se mostrou altamente sensivel a qualquer estimulo externo que poderia, inclusive, provocar a morte dos insetos. A alta mortalidade das ninfas de 5o estadio, que foi da ordem de 30% (Fig. 4), foi tambem observada por outros autores. Barretto et al. (1981) destacaram o alto indice de mortalidade de Dipetalogaster maxima (Uhler, 1894) durante a muda, sobretudo no 5o estadio, mas nao conseguiram explicacao para tal fato. Costa et al. (1987) fizeram observacoes semelhantes sobre as anomalias ocorridas durante a muda e confirmaram a alta taxa de mortalidade do D. maxima nesta fase. Casos de ecdises defeituosas e interrupcao do processo da muda imaginal, elevando a mortalidade, tambem foram registrados por Lent e Valderrama (1977) para tres especies do genero Rhodnius e por Rocha et al. (1994) para R. pictipes. Rabinovich (1972) tambem fez referencia a alta mortalidade do T. infestans no 5o estadio, porem nada comentou a respeito de ecdises anormais. Alguns autores atribuem às condicoes de criacao apossimbioticas a alta mortalidade no 5o estadio e na muda imaginal e atrasos no desenvolvimento (Brecher & Wigglesworth 1944, Geigy et al. 1953, Harington 1960). Segundo Lake e Friend (1968), a ausEncia do microorganismo simbionte Nocardia rhodnii (Erikson, 1935) ocasionaria deficiencia de vitaminas do complexo B em R. prolixus. Nyirady (1973) fez estudos comparativos com T. protracta, T. rubida (Uhler, 1894) e R. prolixus criados de forma convencional e assepticamente, com a finalidade de eliminar a N. rhodnii; acompanhando o desenvolvimento de quatro geracoes de insetos em frascos estereis e alimentados em camundongos germfree verificou que nao houve alteracoes no desenvolvimento dos insetos. No presente trabalho os insetos nao foram criados em condicoes apossimbioticas, ja que os ovos tiveram contato com as fezes e as ninfas provavelmente se contaminaram no momento da eclosao. Como nao foi feito nenhum controle a respeito da contaminacao por simbiontes, e diante da complexidade do tema, tornam-se necessarios estudos adicionais, porem, acreditamos que o excessivo manuseio possa ter exercido alguma influencia. AGRADECIMENTOS Ao Dr. Rodrigo Zeledon, da Universidade da Costa Rica pelos especimes que originaram a colonia de T. nitida; ao tecnico Jose Luis da Costa Giesteira, pelo auxilio no acompanhamento das observacoes e a estagiaria Luciana da Fonseca Silva pelo auxilio na manutencao do insetario. REFERENCIAS Barretto AC, Prata AR, Marsden PD, Cuba CC, Trigueira CP 1981. Aspectos biologicos e criacao em massa de Dipetalogaster maximus (Uhler, 1894) (Triatominae). Rev Inst Med Trop Sao Paulo 23: 18-27. Brasileiro VLF, Perondini ALP 1976. Biologia do Triatoma brasiliensis (Hemiptera, Reduviidae, Triatominae) I. Tempo de succao e replecao de ninfas de 4o e 5o estádio. Rev bras Ent 18: 43-50. Brecher G, Wigglesworth VB 1944. The transmission of Actinomyces rhodnii Erikson in Rhodnius prolixus Stal (Hemiptera) and its influence on the growth of the host. Parasitology 35: 220-224. Carcavallo RU, Martinez AM 1972. Life cycles of some species of Triatoma (Hemiptera: Reduviidae). Can Entomol 104: 699-704. Costa JM, Jurberg J 1990. Estudos bionomicos de Cavernicola lenti Barrett & Arias,1985 (Hemiptera, Reduviidae, Triatominae). Mem Inst Oswaldo Cruz 85: 357-366. Costa JM, Jurberg J, Almeida JR 1986. Estudos bionomicos de Dipetalogaster maximus (Uhler, 1894) (Hemiptera-Triatominae). I - Influencia da dieta sobre o ritmo de postura, viabilidade dos ovos, curva de fertilidade e mortalidade das fEmeas. Mem Inst Oswaldo Cruz 81: 365-380. Costa JM, Jurberg J, Almeida JR 1987. Estudos bionomicos de Dipetalogaster maximus (Uhler, 1894) (Hemiptera-Triatominae). II - InfluEncia da dieta sobre o ciclo biologico e resistEncia ao jejum. Mem Inst Oswaldo Cruz 82: 111-118. Dias E 1952. Doenca de Chagas nas Americas. III - America Central. Rev Bras Malariol D trop 4: 75-84. Dias E 1955. Notas sobre o tempo de evolucao de algumas especies de Triatomineos em laboratorio. Rev Brasil Biol 15: 157-158. Dias E 1956. Observacoes sobre a eliminacao de dejecoes e tempo de succao em alguns triatomineos sul-americanos. Mem Inst Oswaldo Cruz 54: 115-124. Dias E, Chandler AC 1949. Molestias humanas transmitidas por hemipteros sugadores. Mem Inst Oswaldo Cruz 47: 403-422. Diotaiuti L, Dias JCP 1987. Estudo comparativo do ciclo evolutivo de Rhodnius neglectus alimentados em pombos ou camundongos. Rev Soc Bras Med Trop 20: 95-100. Ekkens DB 1981. Nocturnal fligths of Triatoma (Hemiptera: Reduviidae) in Sabino Canyon, Arizona. I Light collections. J Med Entomol 18: 211-227. Freitas MG, Costa JO, Costa HMA 1967. Alguns aspectos da biologia do Rhodnius neglectus Lent, 1954 (Hemiptera, Triatominae) em condicoes de laboratorio. I Evolucao. Arq Esc Vet 19: 81-87. Geigy VR, Halff LA, Kocher V 1953. Untersuchungen uber die physiologischen Beziehungen awischen einen Ubertrager der Chagas - Krankheit Triatoma infestans und dessen Darmsymbionten. Schweizerische Med Wochenschrift 39: 928-930. Gonzalez-Angulo W, Ryckman RE 1967. Epizootiology of Trypanosoma cruzi infections in Triatominae and man on the Yucatan Peninsula of Mexico. J Med Ent 4: 44-47. Harington JS 1960. Studies on Rhodnius prolixus: growth and development of normal and sterile bugs, and the symbiotic relatronship. Parasitology 50: 279-286. Heitzmann-Fontenelle TJ 1983/84. Bionomia comparativa de triatomineos. II - Triatoma brasiliensis Neiva,1911(Hemiptera, Reduviidae). Mem Inst Butantan 47/48: 157-164. Juarez E 1970. Comportamento do Triatoma infestans sob varias condicoes de laboratorio. Rev Saude Publ S Paulo 4: 147-166. Jurberg J, Galvao C, Barth OM 1991. Estudo morfologico de ovos e ninfas de Triatoma nitida Usinger, 1939(Hemiptera, Reduviidae,Triatominae). Rev Brasil Biol 51: 365-372. Kirk ML, Schofield CJ 1987. Density-dependent timing of defaecation by Rhodnius prolixus, and its implications for the transmission of Trypanosoma cruzi. Trans R Soc Trop Med Hyg 81: 348-349. Lafeta LP 1966. Sobre a primeira alimentacao da ninfa de 1 estadio do Panstrongylus megistus Burmeister (1835). Rev Brasil Malariol D trop 18: 113-124. Lake P, Friend WG 1968. The use of artificial diets to determine some of the effects of Nocardia rhodnii on the development of Rhodnius prolixus. J Insect Physiol 14: 543-562. Lehane MJ, Schofield CJ 1981. Field experiments of dispersive flight by Triatoma infestans. Trans R Soc trop Med Hyg 75: 399-400. Lehane MJ, Schofield CJ 1982. Flight initiation in Triatoma infestans (Klug) (Hemiptera: Reduviidae). Bull Ent Res 72: 497-510. Lent H, Jurberg J 1969. Observacoes sobre o ciclo evolutivo, em laboratorio, do Panstrongylus geniculatus (Latreille, 1811) (Hemiptera, Reduviidae, Triatominae). An Acad Brasil CiEnc 41: 125-131. Lent H, Jurberg J 1992. Estudo comparativo da genitalia externa masculina deTriatoma neotomae Neiva, 1911 e Triatoma nitida Usinger, 1939 (Hemiptera, Reduviidae). Mem Inst Oswaldo Cruz 87: 123-130. Lent H, Valderrama A 1977. Observacoes, em laboratorio, sobre o ciclo evolutivo de Rhodnius prolixus Stal, 1859, R. pictipes Stal, 1872 e R. neivai Lent, 1953. Rev Brasil Biol 37: 325-344. Lent H, Wygodzinsky P 1979. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of ChagasÆ disease. Bull Am Mus Nat Hist 163: 123-520. Leon JR 1959. Estado Actual de la Enfermedad de Chagas en Guatemala. Resumen Epidemiologico. Rev Goiana Med 5: 445-455. Leopoldo-e-Silva R, Correa FMA, Curi PP 1990. Influencia da precocidade do primeiro repasto no desenvolvimento pos-embrionerio de Triatoma infestans Klug, 1834 e de Rhodnius prolixus Stal, 1859 (Hemiptera, Reduviidade, Triatominae) em condicoes de laboratorio. Rev bras Ent 34: 361-367. McEwen PK, Lehane MJ 1993. The effect of varying feed interval on nymph development, subsequent adult flight behaviour and autogeny, in the triatominae bug Triatoma infestans (Klug) (Hem., Reduviidae). J Appl Ent 115: 90-96. Nyirady SA 1973. The germfree culture of three species of Triatominae: Triatoma protracta (Uhler); Triatoma rubida (Uhler) and Rhodnius prolixus Stal. J Med Ent 10: 417-448. Ozete HJ 1981. Observaciones sobre la biologia de Triatoma flavida Neiva, 1911 en Cuba. Rev Cub Med Trop 33: 42-50. Perlowagora-Szumlewicz A 1969. Estudos sobre a biologia do T. infestans, o principal vetor da doenca de Chagas no Brasil (Importancia de algumas de suas caracteristicas biologicas no planejamento de esquemas de combate a esse vetor). Rev Brasil Malariol D trop 21: 117-159. Pippin WF 1970. The biology and vector capability of Triatoma sanguisuga texana Usinger and Triatoma gerstaeckeri (Stal) compared with Rhodnius prolixus (Stal) (Hemiptera: Triatominae). J Med Ent 7: 30-45. Rabinovich JE 1972. Vital statistics of Triatominae (Hemiptera: Reduviidae) under laboratory conditions I Triatoma infestans Klug. J Med Ent 9: 351-370. Rocha DS, Galvao C, Jurberg J 1994. Biologia do Rhodnius pictipes Stal, 1872 em condicoes de laboratorio (Hemiptera, Reduviidae, Triatominae). Mem Inst Oswaldo Cruz 89: 265-270. Silva HHG, Silva IG 1991. Ciclo evolutivo de Triatoma dimidiata capitata Usinger, 1944 (Hemiptera, Reduviidae). An Soc Ent Brasil 20: 405-410. Souza JMP, Rodrigues VLCC, Rocha-e-Silva EO 1978. Triatoma sordida - consideracoes sobre o tempo de vida das formas adultas e sobre a oviposicao das femeas. Rev Saude Publ Sao Paulo 12: 291-296. Tolezano JE, Araujo MFS, Chieffi PP, Valentim AM, Ribeiro SS 1984. Variaveis relacionadas ao desenvolvimento de Triatoma infestans Klug, 1834 em condicoes de laboratorio. Efeitos da manipulacao na sobrevivEncia e na fertilidade. Rev Inst Adolfo Lutz 44: 81-86. Wood SF 1951. Importance of feeding and defecation times of insect vectors in transmission of ChagasÆ disease. J econ Ent 44: 52-54. Wood SF 1976. Body weight and blood meal size in Triatoma protracta (Hemiptera: Reduviidae). Ann Entomol Soc Am 69: 632-634. Zárate LG, Lopez GM, Ozuna MC, Santiago GG, Zarate RJ 1984. The biology behaviour of Triatoma barberi (Hemiptera: Reduviidae) in Mexico. IV. Feeding and defecation patterns. J Med Entomol 21: 548-560. Zeledon R, Alvarado R, Jiron LF 1977. Observations on the feeding and defecation patterns of three triatominae species (Hemiptera:Reduviidae). Acta Tropica 34: 65-77.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc95134d.gif] [oc95134b.gif] [oc95134c.gif] [oc95134a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}