 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz Interaction between Didelphis albiventris and Triatoma infestans in Relation to Trypanosoma cruzi Transmission Nicolas J Schweigmann, Silvia Pietrokovsky, Victoria Bottazzi, Osvaldo Conti, Cristina Wisnivesky-Colli/+ Unidad Ecologia de Reservorios y Vectores de Parasitos, Departamento de Ciencias Biologicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 1428, Buenos Aires, Argentina
Code Number: OC95137
Size of Files:
Text:
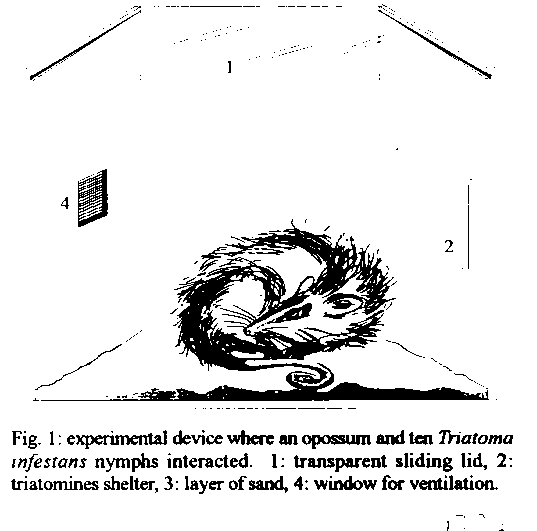
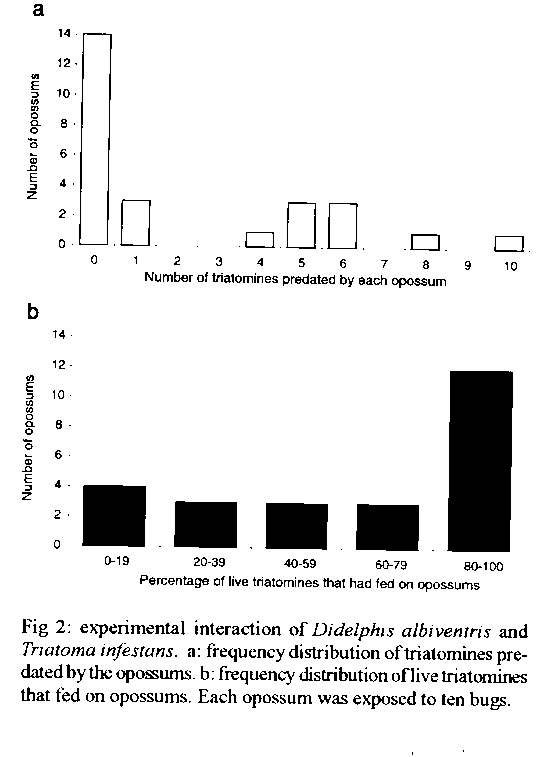
Graphics: Line Drawings (gif) - 16KThis paper attempts to prove if a high Trypanosoma cruzi prevalence of opossums might be reached with few potential infective contacts. One non-infected Didelphis albiventris to T. cruzi and 10 infected nymphs of Triatoma infestans were left together during 23 hr in a device that simulated a natural opossum burrow. Twenty-six replicates were performed using marsupials and triatomines only once. Potentially infective contacts occurred in all the trials. From the 26 opossums used in trials, 54% did not eat any bug. Of the 260 bugs used, 21% were predated. In the 25 trials involving 205 surviving bugs, 36% of them did not feed. In 15/25 cases, >/= 60% of the triatomines were able to feed. The parasitological follow-up of 24 opossums showed that among 10 that had eaten bugs, 4 turned out infected and among the 14 that had not predate, 3 (21%) became positive. In sum, 7/24 (29%) of the marsupials acquired the infection after the experiment. This infection rate was similar to the prevalences found for the opossum population of Santiago del Estero, Argentina, suggesting that the prevalences observed in the field might be reached if each marsupial would encounter infected bugs just once in its lifetime. Key words: opossums - Triatoma infestans - Didelphis albiventris - Trypanosoma cruzi - experimental infection - Argentina Opossums of the genera Didelphis are the most important wild reservoirs of American trypanosomiasis, since their three widely distributed species harbour Trypanosoma cruzi in every ecosystem where they were studied (Barretto & Ribeiro 1979, Schweigmann 1994). These marsupials show omnivorous feeding habits, and insects are also part of their diet (Cordero et al. 1987). Pieces of triatomines were found by Crespo (1982) in the stomach content of one D. marsupialis. Opossums proved to be highly susceptible to the T. cruzi infection by the oral route. Yaeger (1971) found that two out of seven D. virginiana became infected after eating two adults of Rhodnius prolixus each, and Ribeiro et al. (1987) showed that five out of six D. albiventris acquired the infection after they had eaten Triatoma infestans or R. neglectus. On the other hand, we have not found proven evidences of T. cruzi transmission to opossums through the classic vectorial route. But an indirect confirmation of this last would be that sylvatic triatomines usually feed on Didelphidae as it was demonstrated by the finding of opossum blood in the gut contents of bugs collected in wild ecotopes of several countries (Zeledon 1972, Pifano 1973, Christensen & Vazquez 1981, Wisnivesky-Colli et al. 1987). In the province of Santiago del Estero, Argentina, D. albiventris shelters in cavities of trees and cacti, under bromeliad leaves and in burrows in the ground (Schweigmann 1994), where triatomines are also found. The feeding profile of the sampled insects indicated that they all had fed on Didelphidae (Wisnivesky-Colli et al. 1992). This fact would suggest the existence of a trophic interaction between both marsupials and insects, implying the existence of a possible focus of parasite transmission. In the same study area, the opossum population showed T. cruzi annual infection rates from 29 to 50% (Schweigmann 1994). In contrast, less than 1% of the triatomines sampled from opossums shelters (0 to 14 insects per biotope, unpublished results) showed "T. cruzi like" flagellates under microscopical observation (Wisnivesky-Colli et al. 1993). How this high opossum prevalence goes along with such few infected vectors is still unclear. Other trypanosomatids like African trypanosomes and leishmanias modify the feeding behaviour of infected vectors thus increasing the frequency and duration of biting (Lehane 1991). In those circumstances, low infection rates of the vectors produce high prevalences in the mammalian hosts. However, the presence of T. cruzi would not affect the feeding behaviour of infected triatomines (D'Alessandro & Mandel 1969). This high infection rate found in opossums could be explained, for example, if these marsupials were highly susceptible to acquiring the infection. With the aim to clarify this fact, in the present work we let infected triatomines and non-infected opossums interact in a device that attempted to simulate a natural burrow. MATERIALS AND METHODS The experiment was carried out during 1990 in a field laboratory in the locality of Amama, province of Santiago del Estero, Argentina. Opossums and triatomines - After capture, juvenile specimens of D. albiventris were kept in separate cages and fed with dog pellets and water. A parasitological follow-up was done before the experiment in order to select non-infected animals, and after it, to check the acquisition of the parasite. In both instances, each animal was analyzed every 30-45 days by xenodiagnosis (Cerisola et al. 1974) using a box with ten third or fourth instar nymphs of T. infestans. Bugs employed in this test were provided by the insectary of the Servicio Nacional de Chagas, and examined at 30, 60 and 90 days after feeding. An opossum was considered as positive when at least one bug of each box was found infected, and as non-infected, after showing three successive negative xenodiagnoses. Following this criterion, 26 negative opossums were used in the experiment. At that time they already were 6.5 - 9.8 months old (Schweigmann 1994), corresponding to the preadult or adult stages described by Gilmore (1943). In order to work with sylvatic strains of T. cruzi, 260 laboratory bred third and fourth instar nymphs of T. infestans were infected from positive opossums captured in the same study area. These insects were checked out individually to determine the presence of the parasite. Experimental arena - The experimental device (adapted from Rossel 1984) (Fig. 1) consisted of a hardboard box (50 x 50 x 45 cm) - where the opossum could move freely - with a transparent sliding lid at the top. On one of the walls it had a removable wooden panel (40 x 14 cm), which acted as a shelter for triatomines. It was raised 5 cm above the floor and 0.5 cm from the wall, leaving its lateral and lower edges open. A window (10 x 10 cm) covered with fine wire net provided ventilation. A thin layer of sand was spread on the floor to absorb moisture from the excreta. Experimental design - Non-infected opossums were starved for 1-2 days and infected triatomines were fasted for 20-30 days before the trials. Based on the number of triatomines found in the natural biotopes, we decided to use 10 bugs in each experimental test. Insects were introduced first - at approximately 6.30 pm - to allow them to hide, and one hour later one opossum was released into the cage. They were left together for 23 hr, and afterwards the box was opened and examined, recording the number and nutritional status of live bugs. Rossell method (1984) was used to determine if bugs had fed or not, and any nymph with a convex abdomen was regarded as fed. Missing bugs were considered as predated by the opossum. In order not to interfere with the ongoing interaction in the box, no direct observations were made during trials. Twenty-six replicates were done using marsupials and insects only once. RESULTS There have been potentially infective contacts in all the trials, that is, opossums predated on triatomines and/or the bugs fed on the marsupials. The overall proportion of predated bugs was 55/260 (21%). In Fig. 2a is shown the frequency distribution of bugs eaten by opossums. From the 26 opossums used in the trials, 46% ate at least one triatomine, and only one mammal predated all insects. Considering the 25 trials where surviving bugs were found (n=205), 36% of them did not feed. The frequency distribution of live and fed bugs is shown in Fig. 2b. It is noticeable that in 15/25 cases most of the triatomines (=/> 60%) were able to feed. To determine the proportion of opossums that had acquired the infection parasitological follow-ups of 24 marsupials (two animals died in the meantime) were carried out for five months after the tests. Among the 10 opossums that ate bugs, 4 (40%) turned out positive to T. cruzi and among the 14 that did not predate, 3 (21%) became infected. The difference between these percentages was not significant. In brief, a total of 7 (29%) of marsupials got the infection after the experiment. It is worth mentioning two extreme cases: an opossum which predated all bugs and did not become infected and a non-predator that served as blood-source for only one bug but acquired the infection. DISCUSSION The following features make our model realistic: (a) it involves opossums captured in the study area, as well as triatomines infected with T. cruzi strains isolated from these marsupials; (b) the use of nymphs because of their abundance and frequency compared to adults found in opossum burrows (Wisnivesky-Colli 1994); (c) it offers the opossums a place of similar dimensions to their burrows; (d) it provides a shelter for triatomines as opossum burrows do; (e) the number of bugs (10) that interacted with opossums is similar to that found in natural burrows. Simulating a natural ecotope, made it extremely difficult to determine the number, size and distribution of hiding sites for triatomines in the box. So we finally decided on a single refuge that covered 4.5% of the inner surface of walls and floor - the only places where bugs were able to walk. Even if in nature, refuges for triatomines were more abundant, under our experimental conditions it was observed that the number of insects that could feed avoiding predation was high. This is reinforced by the fact that most of opossums ate 0 - 1 bug. However, it may be possible that bugs do not constitute an attractive meal for opossums. This result may suggest that most of the triatomines actively searched for food during the sleeping periods of opossums so that their risk of death was minimized; we cannot disregard the possibility that many bugs may have been predated because of the distress they had provoked on the half asleep opossum while biting. Hunsaker (1977) stated that the metabolic activity of opossums diminishes when they sleep. Therefore, it is expected that opossums are less responsive than eutherians while resting, and this fact would account for both the low predation and the numerous fed bugs recorded at the end of the experiment. It is worthwhile mentioning that during the experiment the marsupials did not show behavioural alterations related to stress. Opossums show crepuscular and nocturnal activity and rest in their burrows during the day (Hunsaker 1977). Therefore, it should be expected the oral route to represent the main way in of parasites during the activity periods while the classic vectorial route would prevail during daytime. In this experiment, opossums that ate bugs might have acquired the infection by either the vectorial and/or the oral routes; those that did not predate, could have become infected by the vectorial route or by licking contaminated faeces. To actually determine this last point, it is necessary to perform other experiments. Although the percentage of infected opossums that had eaten bugs (40%) was higher than that showed by those that had not eaten (21%), the difference was not significant. However, predators showed a trend to a higher probability of acquiring the infection as they would be exposed to a double transmission route: vectorial and/or oral. Due to the opossums seminomadic habits (Hunsaker 1977) they change of burrows periodically and so they are frequently exposed to the triatomines that inhabit the different shelters. The number of bugs present in each burrow ranges from 0 - 14 with a very low rate of infection. Therefore, it is reasonable to consider that an adecuate number of potentially infective contacts would be ten for the 1-1.5 years of the opossum's lifespan (Schweigmann 1994). We decided to expose the marsupials to the risk they would face in their entire lifetime but within 23 hr. Rabinovich et al (1990), estimated that about 1,500-2,000 infective contacts are needed to infect a child under 15 years old in the same endemic area we are studying. Since in our experiment opossums became infected with few potential infective contacts (1 to 10) in a short lapse, we may assume that opossums are more susceptible than humans to the infection. The percentage of infection (29%) was similar to the prevalence recorded in the population of opossums. Consequently, we think that the high opossum prevalences observed in the field might be reached if each marsupial would encounter just once in its lifetime with infected bugs. Further research is needed to prove this assumption.
ACKNOWLEDGEMENTS To the Citatti family for providing the animal house at the field, to the Moyanos for hostage, to Bea Ferrer for her kind assistance with the English version of the manuscript, and to Mario Capovia for his drawing. REFERENCES Barretto MP, Ribeiro RD 1979. Reservatorios silvestres do Trypanosoma cruzi. Rev Inst Adolfo Lutz 39: 25-26. Cerisola JA, Rohwedder R, Segura EL, Del Prado CE, Alvarez M, Wynne de Martini GL 1974. El xenodiagnóstico. Ministerio de Bienestar Social, Buenos Aires, Argentina, 127 pp. Cordero GA, Ruben A, Nicolas B 1987. Feeding Habits of the Opossum (Didelphis marsupialis) in Northern Venezuela. Fieldiana: Zoology 39: 125- 131. Crespo JA 1982. Ecología de la comunidad de mamíferos del Parque Nacional Iguazú, Misiones. Museo Argentino de Ciencias Naturales B Rivadavia. Ecología. Tomo III, 162 pp. Christensen HA, Vazquez A 1981. Host feeding profiles of Rhodnius pallecens. Am J Trop Med Hyg 30: 278-283. D'Alessandro A, Mandel S 1969. Natural infections and behaviour of Trypanosoma rangeli and Trypano-soma cruzi in the vector Rhodnius prolixus in Colombia. J Parasitol 54: 846-852. Gilmore RM 1943. Mammalogy in an epidemiological study of jungle yellow fever in Brazil. J Mamm 24: 144-162. Hunsaker D 1977. Ecology of New World Marsupials, p. 95-156. In D Hunsaker, The Biology of Marsupials. II Ed. Academic Press, NY, U.S.A. Lehane MJ 1991. Biology of blood-sucking insects. HarperCollins Academic, Chapman & Hall, Hampshire, England, 279 pp. Pifano FC 1973. La dinámica de la enfermedad de Chagas en el Valle de los Naranjos. Estado de Carabobo. Venezuela. Arch Venez Med Trop Parasitol Med 5: 1-29. Rabinovich JE, Wisnivesky-Colli C, Solarz ND, Gurtler RE 1990. Probability of transmission of Chagas disease by Triatoma infestans (Hemiptera: Redu-viidae) in an endemic area of Santiago del Estero, Argentina. Bull WHO 68: 737-746. Ribeiro RD, Rissato e Garcia TA, Bonoma WCh 1987. Contribuição para o estudo dos mecanismos de transmissão do agente etiologico da doenca de Chagas. Rev Saude Publ 21: 51-54. Rossell O 1984. A comparison of the feeding performances of Rhodnius prolixus Stal and Rhodnius robustus Larrousse (Hemiptera: Triatominae). Doctoral Thesis, University of London, 339 pp. Schweigmann NJ 1994. Aspectos ecologicos de una poblacion santiaguena de la comadreja overa (Didelphis albiventris) en relacion con la transmision del Trypanosoma cruzi. Doctoral Thesis, Universidad de Buenos Aires, 184 pp. Wisnivesky-Colli C 1994. Triatominos vectores secundarios de Trypanosoma cruzi: su domicilia-cion potencial. Taller Latinoamericano de Control de Vectores, Venezuela, 3: 83-89. Wisnivesky-Colli C, Ruiz A, Ledesma OAM, Gurtler RE, Lauricella M, Salomon DO, Solarz ND, Segura EL 1987. Ecologia domestica de la tripanosomiasis americana: Perfil alimentario de Triatoma infestans en un area rural de Santiago del Estero. Rev Soc Bras Med Trop 20: 31-39. Wisnivesky-Colli C, Schweigmann NJ, Alberti A, Pietrokovsky S, Conti O, Montoya S, Riarte A, Rivas C 1992. Sylvatic American Trypanosomiasis in Argentina. Trypanosoma cruzi infection in mammals from the Chaco Forest in Santiago del Estero. Trans R Soc Trop Med Hyg 86: 38-41. Wisnivesky-Colli C, Schweigmann N, Pietrokovsky S, Bottazzi V, Conti O 1993. Pecularities of sylvatic transmission of Trypanosoma cruzi in northern Argentina. Mem Inst Oswaldo Cruz 88 (Suppl) p. 266, VE-35. Yaeger RG 1971. Transmission of Trypanosoma cruzi infection to opossums via the oral route. J Parasitol 57: 1375-1376. Zeledon R 1972. Epidemiology, modes of transmission and reservoir hosts of Chagas disease. Ciba Foundation, Symposium 20, Elsevier Excerpta Medica, Amsterdam: 57-77. This research work was performed with financial support from Secretaria de Ciencia y Tecnica, Universidad de Buenos Aires, Argentina. +Corresponding author Received 28 December 1994 Accepted 13 June 1995 Copyright 1995 Fundacao Oswaldo Cruz (FIOCRUZ) The following images related to this document are available:Line drawing images[oc95137a.gif] [oc95137b.gif] |
| |||||||||

{kind=link}
{kind=link}