 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz Neotropical Monogenoidea. 24. Rhinoxenus bulbovaginatus n. sp. (Dactylogyridae, Ancyrocephalinae) from the Nasal Cavity of Salminus maxillosus (Osteichthyes, Characidae) from the Rio Parana, Parana, Brazil Walter A Boeger, Marcus V Domingues, Gilberto C Pavanelli* Departamento de Zoologia, Universidade Federal do Parana, 81531-990 Curitiba, PR, Brasil *NUPELIA/DBI, Universidade Estadual de Maringa, 87020-900 Maringa, PR, Brasil
Code Number: OC95141
Size of Files:
Text:
Graphics: Line Drawings (gif) - 33KRhinoxenus bulbovaginatus n. sp. is described from the nose of Salminus maxillosus (Characidae) collected in the basin of the Rio Parana, near the city of Porto Rico, State of Parana, Brazil. The new species can be differentiated from the other three species in the genus by the morphology of the copulatory complex, vagina, and ventral anchor. The sister group relationship of the known species of Rhinoxenus was determined using techniques of Phylogenetic Systematics (Cladism). The resulting cladogram (C.I.=100%) indicates that the new species is most closely related to R. piranhus Kritsky, Boeger and Thatcher, 1988. The other two species of the genus, R. arietinus Kritsky, Boeger and Thatcher, 1988 and R. nyttus Kritsky, Boeger and Thatcher, 1988, both parasites of Anostomidae fishes, have a paraphyletic position in the cladogram, suggesting that the origin of at least one of them can not be associated to cospeciation. Key words: Dactylogyridae - Ancyrocephalinae - Rhinoxenus bulbovaginatus n. sp. - Salminus maxillosus - phylogeny Only two genera of Dactylogyridae are known to include species from the nasal cavities of Characiformes fishes of the Neotropical region: Rhinonastes Kritsky, Thatcher and Boeger, 1988, and Rhinoxenus Kritsky, Boeger and Thatcher, 1988. Rhinonastes pseudocapsaloideum, the type and only species of the genus, parasitizes prochilodontid characins (Kritsky et al. 1988a). All known species of Rhinoxenus occur on members of two characin families. Rhinoxenus arietinus and R. nyttus parasitize Anostomidae fishes while R. piranhus is found in Serrasalmidae. During a survey of nose parasites of fishes from the Parana River, a new species of Rhinoxenus was collected from the "dourado", Salminus maxillosus Valenciennes, 1849, a Characidae. This species is described herein and its phylogenetic position within the genus determined using techniques of Phylogenetic Systematics.
MATERIALS AND METHODS
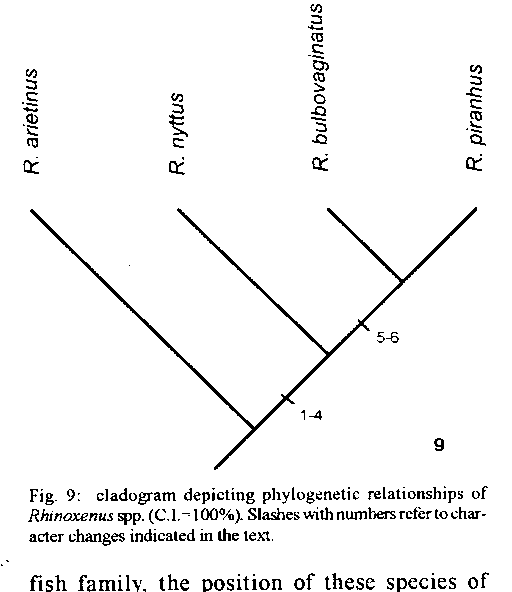
Hosts (S. maxillosus) were collected by the personnel of the NUPELIA, Universidade of Maringá, in the basin of the Rio Parana near the city of Porto Rico, State of Parana, Brazil. Methods of parasite collection, preparation, measurement, and illustration are as described by Kritsky et al. (1986). Measurements are in micrometers, the mean is followed by the range in parentheses. Type specimens are deposited in helminthological collections of the Instituto Oswaldo Cruz (IOC), Rio de Janeiro, Brazil; U.S. National Museum (USNM), Beltsville, Maryland, and the University of Nebraska State Museum (HWML) Lincoln, Nebraska. Transformation series used in the phylogenetic analysis were defined from the literature and available specimens. Homologous series in which apomorphic state represents an antapomorphy of a single ingroup taxon were not utilized. Ingroup taxa included all known species of Rhinoxenus - R. piranhus, R. arietinus, R. nyttus, all described by Kritsky et al. (1988a) and the new species described herein. The hypothesis on the evolutionary relationship of the species of Rhinoxenus was constructed using Hennigian Argumentation (Wiley 1981); the topology of the resulting cladogram was then subjected to PAUP (Phylogenetic Analysis Using Parsimony, Version 2.4.1; D L Swofford, Smithsonian Institution, Washington, DC) to confirm that it was among the most parsimonious trees. A total of 11 character states comprising six transformation series was utilized in the analysis. Polarization of homologous series was determined by outgroup comparison (Watrous & Wheeler 1981). Amphithecium Boeger and Kritsky, 1988 and Rhinonastes Kritsky, Thatcher and Boeger, 1988 were utilized as outgroups. Homologous series follow numbers in parentheses, preceeding the definition of a character state, refer to the coding that state received in the data matrix (Table); numbers in brackets, following the definition, refer to respective evolutionary changes depicted in the cladogram (Fig. 9). 1. Eyes: plesiomorphy - (0) pairs 4 equidistant, apomorphy - (1) members of posterior pair closer together than members of anterior pair [5]; 2. Ventral anchor: plesiomorphy - (0) basal protuberance at articulation of bar absent, apomorphy - (1) protuberance at articulation of bar present [l]; 3. Point of ventral anchor: plesiomorphy - (0) almost as long as shaft, apomorphy - (1) shorter than shaft [6]; 4. Base of ventral anchor: plesiomorphy - (0) superficial and deep roots well developed, apomorphy - (1) superficial and deep roots inconspicuous [2]; 5. Bar: plesiomorphy - (0) with slightly expanded end, apomorphy - (1) with protuberance for articulation of ventral anchor [3]; 6. Hook pair 2: plesiomorphy - (0) at level of germarium, apomorphy - (1) posterior to level of germarium. TABLE
Matrix of characters of Rhinoxenus spp --------------------------------------- R. arietinus 000000 R. piranhus 111111 R. nyttus 010111 R. bulbovaginatus 111111 Ancestor 000000 DESCRIPTION
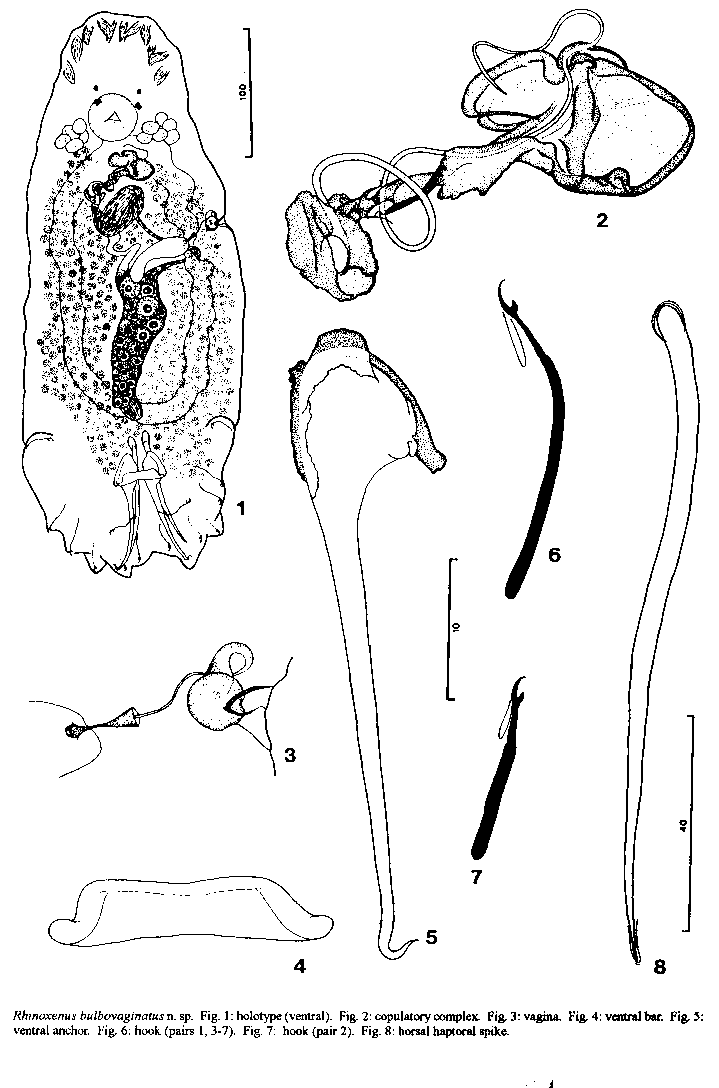
Description (based on 20 specimens): body 441 (430-454; n=4) long, flattened dorsoventrally; greatest width 160 (115-190; n=4) at body midlength. Cephalic area with poorly developed terminal lobes; head organs conspicuous, usually 4 pairs; cephalic glands conspicuous, unicellular, posterolateral to pharynx. Eyes usually equidistant, members of anterior pair smaller than those of posterior pair; eye granules ovate, small; accessory granules not observed. Pharynx spherical, 41-43 (n=2) in diameter. Haptor trapezoidal, 100-105 (n=2) long, 102-110 (n=2) wide. Ventral anchor 113 (110-119; n=8) long, with undifferentiated superficial and deep roots, base with protuberance serving as articulation for bar, straight shaft, fish-hook-shaped point; base 26 (23-29, n=9) wide. Dorsal haptoral spike 119 (115-122; n=5) long, with bulbous proximal end, tapered shaft, pointed distal end with conspicuous cap of tissue. Ventral bar trapezoidal, with short posterolateral protuberances, anterolateral margin heavily sclerotized; bar 42 (35-54- n=8) long. Hook pair 2, 31 (29-33; n=3) long, lying on 2 bilateral lobes on posterior trunk, erect thumb; proximal 2/3 of shank inflated; other hooks 41 (29-52; n=16) long, elongate, with curved point, erect thumb, shank sligthly inflated proximally, FH loop extending to near beginning of dilated portion of shank. Copulatory organ a coil of about 1½ rings, with wide, flat sclerotized base. Accessory piece articulated to base of copulatory organ, with expanded distal end. Testis elongate, 95 (81-118; n=3) long, 24 (20-27; n=3) wide; seminal vesicle fusiform, without median constriction; prostatic reservoirs not observed. Germarium elongate, 82-102 (n=2) long, 27-32 (n=2) wide; ootype, uterus not observed. Vagina sclerotized, connected to seminal receptacle through a sclerotized cone-shaped duct, widening distally, with distal loop and bulb; vaginal aperture wide, with sclerotized walls. Vitellaria dense, coextensive with gut. REMARKS Rhinoxenus bulbovaginatus n. sp. is morphologically distinct from the previoulsy described species of the genus based mainly on the morphology of the copulatory complex, vagina, and ventral anchor. It represents the first species of the genus described and/or reported from outside the Amazon and on a Characidae. The specific name refers to the presence of a bulb in the vagina of this species. The cladogram (C.I. = l00 %), depicting the sister group relationship of species of Rhinoxenus, is presented in Fig. 9. Rhinoxenus bulbovaginatus is the sister species of R. piranhus with which it shares a reduced point on the ventral anchor, eyes equidistant, and point of ventral anchor shorter than shaft. The other two species of the genus, R. arietinus and R. nyttus, are both parasites of Anostomidae fishes, but do not depict an exclusive single ancestor. Assuming monophyli of this fish family, the position of these species of Rhinoxenus in the cladogram suggests that the origin of at least one of them can not be associated with cospeciation. ACKNOWLEDGEMENTS To Dr Delane C Kritsky (ldaho State University, USA) for providing a presubmission revision of the manuscript. REFERENCES Kritsky DC, Boeger WA, Thatcher VE 1988a. Neotropical Monogenea. 11. Rhinoxenus, new genus (Dactylogyridae, Ancyrocephalinae) with descriptions of three new species from the nasal cavities of Amazonian Characoidea. Proc helm Soc Wash 101: 87-94. Kritsky DC, Thatcher VE, Boeger WA 1988b. Neotropical Monogenea. 13. Rhinonastes pseudo-capsaloideum n.gen., n.sp. (Dactylogyridae, Ancyrocephalinae), a nasal parasite of curimatã, Prochilodus nigricans Agassiz (Cypriniformes, Prochilodontidae), in Brazil. J Parasit 74: 695-698. Kritsky DC, Thatcher VE , Boeger WA 1986. Neotropical Monogenea. 8. Revision of Urocleidoides (Dactylogyridae, Ancyrocephalidae). Proc helm Soc Wash 53: 1 37. Watrous LE, Wheeler QD 1981. The outgroup comparison method of character analysis. Syst Zool 30: 1-11. Wiley EO 1981. Phylogenetics. The Theory and Practice of Phylogenetic Systematics. John Wiley and Sons, New York, 439 pp. This work received financial support from the Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq). This is the contribution number 883 from the Departamento de Zoologia, Universidade Federal do Parana. Research fellows CNPq Received 13 December 1994 Accepted 12 April 1995 Copyright 1995 Fundacao Oswaldo Cruz, FIOCRUZ
The following images related to this document are available:Line drawing images[oc95141a.gif] [oc95141b.gif] |
| |||||||||

{kind=link}
{kind=link}