 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz Vol. 90(6), Nov./Dec. 1995 Natural Hosts of Notocotylus breviserialis (Digenea, Notocotylidae) Parasite of Brazilian Waterfowl Luis Claudio Muniz-Pereira+, Suzana B Amato++ Departamento de Biologia Animal, Universidade Federal Rural do Rio de Janeiro, Caixa Postal 74512, 23851-970 Seropedica, RJ, Brasil
Code Number: OC95145
Size of Files:
Text:
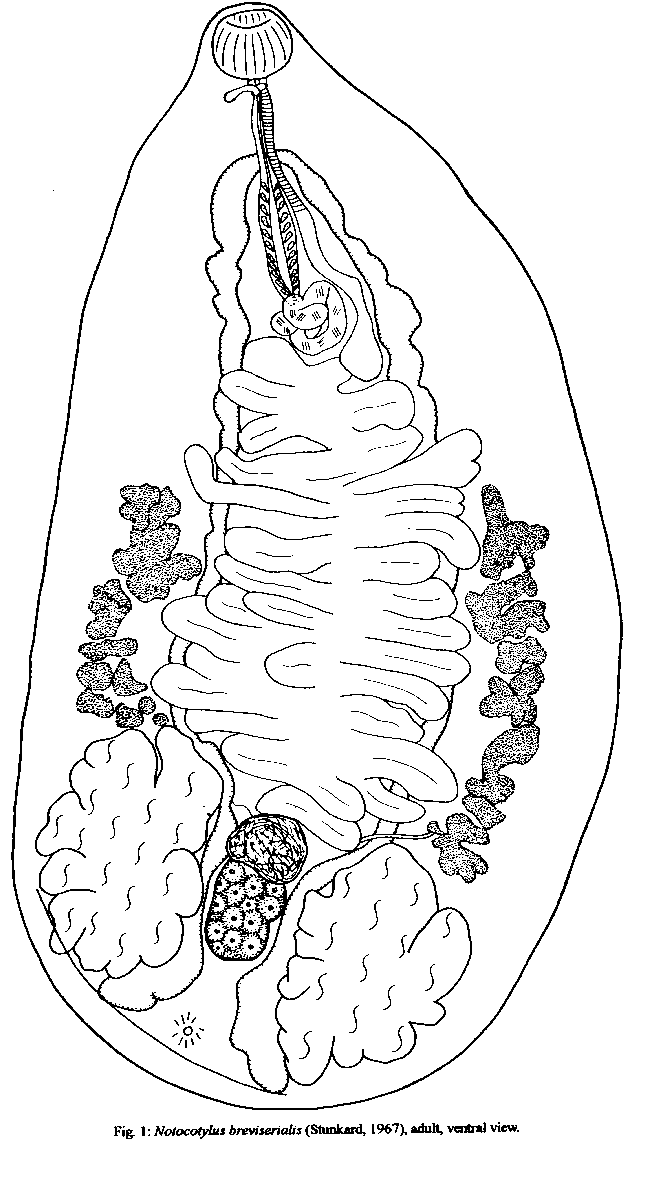
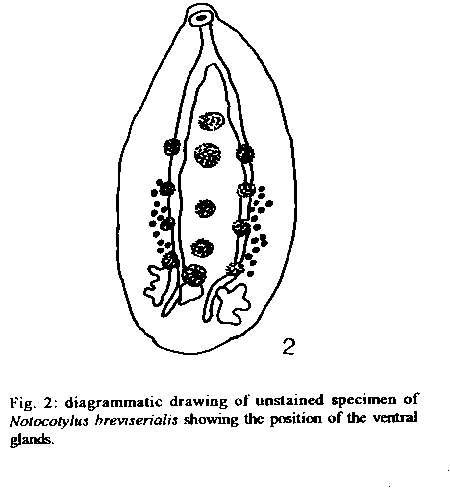
Graphics: Line Drawings (gif) - 32KNotocotylus breviserialis has been collected from Anas bahamenis L. and Amazonetta brasiliensis (Gmelin), in Marica, State of Rio de Janeiro, Brazil. It is the first time that this digenean is collected as adult in its natural hosts, and this is its first record for the Neotropical region. The prevalence of N. breviserialis was 5.56% and 18.18% in A. bahamensis and in A. brasiliensis, respectively. New morphological data about the location of the vitelline glands and previtellinic space measurements are presented. Key words: Notocotylus breviserialis - Notocotylus - Digenea - Anatidae - Anas - Amazonetta - natural host - Neotropical region - Brazil Notocotylus breviserialis, a parasite of the bursa of Fabricius, was described by Stunkard (1967), based on specimens recovered from experimentally infected domestic ducks, Anas platyrhynchos L., and chickens, Gallus gallus L. Beverley-Burton (1958) originally allocated N. breviserialis in the genus Uniserialis, when U. gippyensis was described. This genus was posteriorly synonimyzed by Baer and Joyeaux (1961) with Notocotylus Diesing, 1839. Bisset (1977) proposed the new combinations N. breviserialis and N. gippyensis. MATERIALS AND METHODS A total of 18 Anas bahamensis and 22 Amazonetta brasiliensis were collected, with license from the Instituto Brasileiro do Meio Ambiente (IBAMA), between November 1989 and January 1992, from the Marica and Guarapina lagoons and adjoining mudflat areas, in Marica District, State of Rio de Janeiro, Brazil. Host specimens were placed on ice, in individual plastic bags, for a few hours and were later frozen to be examined in the next morning. Their sex, weight, total length, and wing length were recorded. The birds are deposited in the Ornithological Collection of the Museu Nacional do Rio de Janeiro, Rio de Janeiro, RJ. During necropsy, all organs were examined for helminths, which were collected, fixed, and prepared as described by Amato et al. (1991). The ecological terms - prevalence, mean intensity of infection, and site of infection - are defined according to Margolis et al. (1982). The measurements given in the description of the specimens are in micrometers, unless otherwise specified; the number inside parentheses is the mean value and ono is the number of specimens measured. Figures were drawn with the aid of a drawing tube on a Leitz Dialux 20EB light microscope. Voucher specimens were deposited in the Colecao Helmintologica, Instituto Oswaldo Cruz (CHIOC), Rio de Janeiro, RJ, Brazil. DESCRIPTION Notocotylus breviserialis (Stunkard, 1967) DESCRIPTION (based on 21 specimens mounted in toto, 7 measured): Notocotylidae, body ovoid, coral pink when alive, 1.72-2.47 mm (2.13 mm) long, 0.933-2.23 mm (1.11 mm) wide, dorsally convex, ventrally concave, anterior extremity atenuatted, posterior rounded (Fig. 1). Ventral surface spined; protusible glands in three rows (Fig. 2)median row with five glands, lateral rows with four glands each, in the intervals between the median row glands. Monostomes. Oral sucker globular, subterminal, 110-168 (153) long, 117-212 (175) wide; pharynx absent; esophagus nearly same length of oral sucker, 153-219 (176) long; ceca long, shape irregular, approximating at ovarian region. Testes lobed, extracecal, left 212-380 (293) long, 139-350 (205) wide, right 241-350 (289, n=6) long, 146-277 (181) wide; vas eferens ventral to ceca; vas deferens dorsal to uterine loops; external vesicle coiled; cirrus pouch 343-599 (460) long, 18-25% of body length; genital pore ventral, posterior to oral sucker, anterior to cecal bifurcation. Ovary lobed, median, intercecal, intertesticular, 95-204 (141) long, 95-168 (133) wide; Mehlis' gland anterior to ovary; uterus intercecal, 12-17 loops, in the second third of body; metraterm thin walled, parallel or ventral to cirrus pouch, opening into genital atrium; follicular vitelline glands, extracecal, 467-732 (557) long, previtellinic space 43-51% of body length; vitellinic ducts ventral to ceca; vitellinic reservoir 27 (n=1) long, 29 (n=1) wide; eggs 18-22 (20) long, 11-13 (12) wide, polar filaments 47-92 long. Excretory pore subterminal, excretory vesicle saccate. Taxonomic summary
Hosts: Anas bahamensis Linne, 1758, Amazonetta brasiliensis (Gmelin, 1758) Site of infection: bursa of Fabricius Locality: mouth of the Mombuca river, Lagoon of Marica, Marica District, State of Rio de Janeiro, Brazil Prevalence: Anas bahamensis 5.56%, Amazonetta brasiliensis 18.18% Mean intensity of infection: Anas bahamensis 8, Amazonetta brasiliensis 6.5 Intensity of infection range: Anas bahamenis 8, Amazonetta brasiliensis 1-17 Other hosts: Aves, Anseriformes: Anas platyrhynchos L. (dom.) [exp.]. Galliformes: Gallus gallus L. (dom.) [exp.] Geographical distribution: South America (Brazil), North America (USA) Specimens deposited: Voucher specimens - CHIOC No. 33287 and 33288 REMARKS This is the first record of a species of the genus Notocotylus in Brazil. Since the description by Stunkard (1967) based on experimental infections of mallard ducks, A. platyrhynchos, and chickens, G. gallus, N. breviserialis was never found in the natural definitive host. The finding of this parasite in Brazil also represents a new geographical record for the Neotropical region. The resemblance of N. breviserialis and N. gippyensis is not so obvious as Bisset (1977) pointed out. In addition to the major morphological differences between these species mentioned by Bisset (1977) we add: (l) the vitelline glands in N. breviserialis are mainly located in the second third of the body length, while in N. gippyensis they are mainly located in the last third; (2) the previtellinic space measured in the drawing of the holotype of N. breviserialis occupies 43% of the body length, while in the drawing of the holotype of N. gippyensis it represented 54%, and in the specimens of the present work it represented 43-51%, but never reaching 54%; (3) in N. breviserialis (present work) the uterine loops occupy intirely the second third of the body length; (4) the position of the genital pore in N. gippyensis is more anterior than it is in N. breviserialis. Notocotylus breviserialis is also similar to N. skrjabini Ablasov (1953), which has as usual site of infection the bursa of Fabricius. Nocotylus skrjabini differs mainly from N. breviserialis by the position of the genital pore, that in the former is located posteriorly to the cecal bifurcation. The disposition of the ventral glands is different in N. skjarbini, having five lateral ventral glands, disposed side by side with the median ones, while in N. breviserialis (Fig. 2) the lateral glands are in the interval of the median ones. These notocotylids have in common hydrobiid snails as intermediate host and species of anatids as definitive hosts. Nocotylus gippyensis was originally described in England and later recorded in New Zealand. The snail intermediate host, Potamopyrgus antipodarum, may have been introduced in England from New Zealand, or possibly Australia, in the 19th century and the anatid host may have been carried to England for ornamental purposes (Bisset 1977). Therefore, this parasite may have been introduced in England from New Zealand. In Brazil, at the study area, we found shells of hydrobiid snails in the stomach contents of the necropsied anatids and Stunkard (1967) found N. breviserialis in hydrobiid snails in the USA. This digenean may have been carried to the region of Maricá by migratory birds, during migration from North to South America.
ACKNOWLEDGMENTS
To Prof. Jorge Bruno Nacinovic and to Prof. Dante M Teixeira, Seção de Ornitologia, Museu Nacional do Rio de Janeiro, Universidade Federal do Rio de Janeiro, for the capture and identification of the anatids and for accepting the anatids for deposit in the Museum's Ornithological Collection, respectively. To Prof. Ricardo M Takemoto, NUPELIA, Universidade Estadual de Maringá, Maringá, PR, for kindly inking the drawing. REFERENCES Ablasov NA 1953. A new trematode of ducks, Notocotylus skrjabini n. sp. Sbornik Rabot po Gelmintologi 75: 15-16. Amato JFR, Boeger WA, Amato SB 1991. Protocolos para laboratorio - coleta e processamento de parasitos do pescado. Imprensa Universitaria, Universidade Federal Rural do Rio de Janeiro, Itaguai, RJ, Brasil, 81 pp. Baer JG, Joyeux C 1961. Classe des trematodes, p. 561-692. In PP Grasse, Traite de Zoologie. Tomo 4, Masson et Cie, Paris, France. Beverley-Burton M 1958. A new notocotylid trematode, Uniserialis gippyensis gen. et sp. nov., from the mallard, Anas platyrhyncha platyrhyncha L. J Parasitol 44: 412-415. Bisset SA 1977. Notocotylus tadornae n. sp. and Notocotylus gippyensis (Beverley-Burton) (Trematoda: Notocotylidae) from waterfowl in New Zealand: morphology, life history and systematic relations. J Helminthol 51: 365-372. Margolis LG, Esch G, Holmes JC, Kuris AM, Schad GA 1982. The use of ecological terms in Parasitology (report of an ad hoc committee of the American Society of Parasitologists). J Parasitol 68: 131-133. Stunkard HW 1967. The morphology, life-history, and systematic relations of the digenetic trematode, Uniserialis breviserialis sp. nov. (Notocotylidae), a parasite of the bursa Fabricius of bird. Biol Bull 132: 266-276. +Present address: Universidade Castelo Branco, Av. Santa Cruz 1631, 21710-250 Rio de Janeiro, RJ, Brasil ++Research Fellow of CNPq Received 2 March 1995 Accepted 26 July 1995 Copyright 1995 Fundacao Oswaldo Cruz, FIOCRUZ
The following images related to this document are available:Line drawing images[oc95145b.gif] [oc95145a.gif] |
| |||||||||

{kind=link}
{kind=link}