 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz Vol. 90(6), Nov./Dec. 1995 Identification of Species Related to Anopheles (Nyssorhynchus) albitarsis by Random Amplified Polymorphic DNA-Polymerase Chain Reaction (Diptera: Culicidae) Richard C Wilkerson/+, Thomas V Gaffigan, Joso Bento Lima* Department of Entomology, Walter Reed Army Institute of Research, Washington, DC 20307 U.S.A. *U.S. Army Medical Research Unit - Brazil, Instituto de Biologia do Exorcito, Rua Francisco Manuel 102, 20911-270 Rio de Janeiro, RJ, Brasil
Code Number: OC95147
Size of Files:
Text:
Graphics: Line Drawing (gif) - 13K
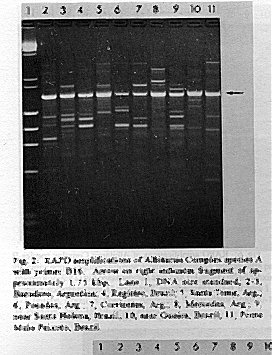
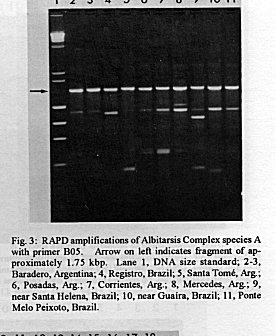
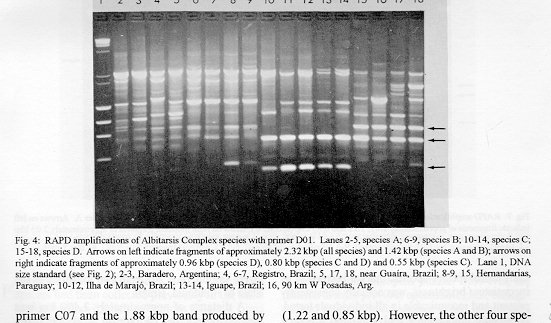
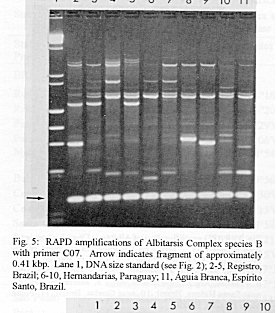
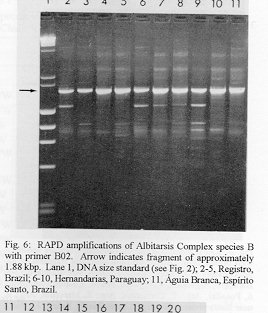
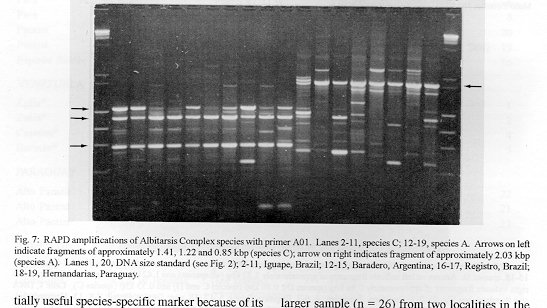
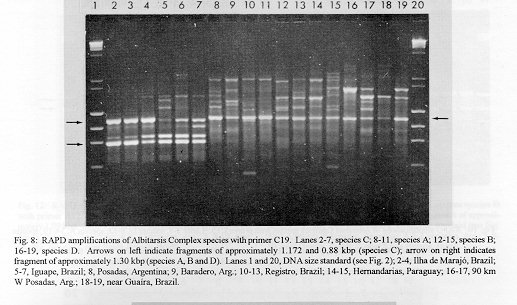
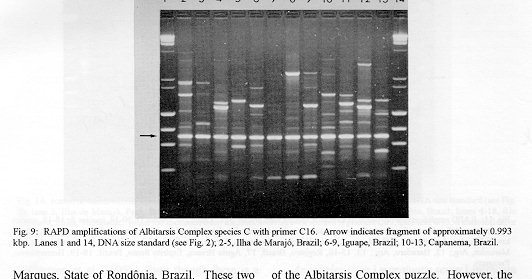
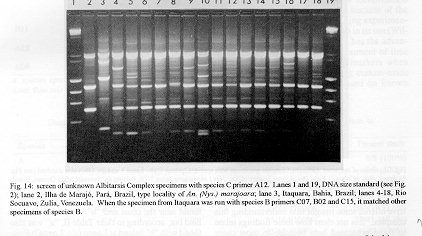
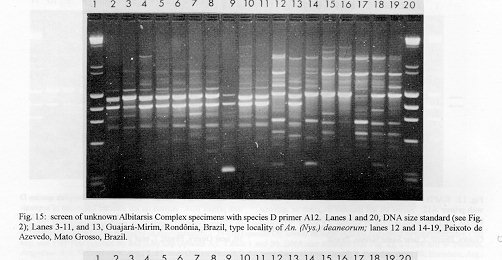
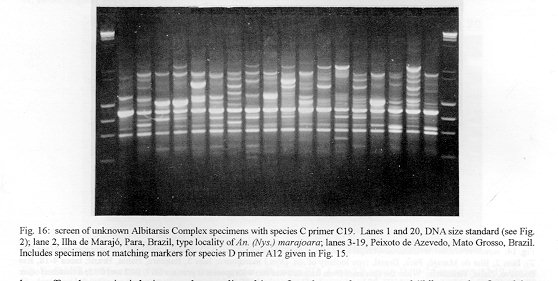
Photographs (jpg) - 443K[NOTE: All figures may be accessed from references at the end of the text.] Species-specific Random Amplified Polymorphic DNA-Polymerase Chain Reaction (RAPD-PCR) markers were used to identify four species related to Anopheles (Nyssorhynchus) albitarsis Lynch-Arribalphalzaga from 12 sites in Brazil and 4 in Venezuela. In a previous study (Wilkerson et al. 1995), which included sites in Paraguay and Argentina, these four species were designated "A", "B", "C" and "D". It was hypothesized that species A is An. (Nys.) albitarsis, species B is undescribed, species C is An. (Nys.) marajoara Galvpio and Damasceno and species D is An. (Nys.) deaneorum Rosa-Freitas. Species D, previously characterized by RAPD-PCR from a small sample from northern Argentina and southern Brazil, is reported here from the type locality of An. (Nys.) deaneorum, Guajarbeta-Mirim, State of Rondônia, Brazil. Species C and D were found by RAPD-PCR to be sympatric at Costa Marques, State of Rondonia, Brazil. Species A and C have yet to be encountered at the same locality. The RAPD markers for species C were found to be conserved over 4,620 km; from Iguape, State of Spio Paulo, Brazil to Rio Socuavo, State of Zulia, Venezuela. RAPD-PCR was determined to be an effective means for the identification of unknown specimens within this species complex. Key words: Anopheles (Nyssorhynchus) albitarsis -Anopheles (Nyssorhynchus) marajoara - Anopheles (Nyssorhynchus) deaneaorum - RAPD PCR - identification The Albitarsis Complex of Anopheles subgenus Nyssorhynchus (Albitarsis Subgroup of Linthicum 1988) is known to include a number of cryptic species (Kreutzer et al. 1976, Steiner et al. 1982, Rosa-Freitas et al. 1990, Narang et al. 1993). Most recently, using Random Amplified Polymorphic DNA - Polymerase Chain Reaction (RAPD-PCR), Wilkerson et al. (1995) demonstrated the existence of four species in the Albitarsis Complex in Brazil, Argentina and Paraguay. Members of this complex have been incriminated as important vectors of malaria parasites (reviewed by Rosa-Freitas et al. 1990) but, because members of the complex are difficult to identify by morphology, it is not known which species might be responsible for transmission in a given area. Our objective was to identify members of the Albitarsis Complex by RAPD-PCR from additional localities and obtain reared associated sibling voucher specimens for furture morphological studies. Becuase of its simplicity, the use of morphological characters is desirable as an aid to research in vector biology, biogeography, malaria epidemiology, malaria drug and vaccine trials and ultimately develop methodologies and strategies for malaria control. In addition, RAPD-PCR was investigated as a possible means for routine identifications in lieu of morphological identification keys. At present, there are three valid species in the complex: An. (Nys.) albitarsis Lynch-Arribbetalzaga, 1878 (Paraguay, southern Brazil and Argentina); An. (Nys.) deaneorum Rosa-Freitas, 1989 (State of Rondônia, Brazil); and An. (Nys.) marajoara Galvpio and Damasceno, 1942 (Costa Rica to Bolivia). Wilkerson et al. (1995) designated the taxa, characterized by RAPD-PCR, as species A, B, C and D. They hypothesized that species A is An. (Nys.) albitarsis sensu stricto, species B is undescribed, species C is An. (Nys.) marajoara and species D is An. (Nys.) deaneorum. RAPD genetic markers found by Wilkerson et al. (1995) were used to identify specimens collected at additional sites in Brazil and Venezuela. A summary of the distribution of the species and photo documentation of these markers is presented (Figs 1-16; Tables I-III). RAPD (Williams et al. 1990, Welsh & McClelland 1990) is a PCR-based technique which has proven to be a quick and effective means of identifying genetic markers. This technique permits numerous markers to be assayed on DNA extracted from a single mosquito. Instead of using primer pairs as in traditional PCR, RAPD reactions use a single short primer (usually 10 bases in length) of randomly chosen sequence. For a RAPD band to be produced, the primer needs to match a binding site that is within approximately 2-3 kilobase pairs of another oppositely oriented binding site so that the single oligonucleotide can prime replication in both the forward and reverse directions. A typical RAPD reaction produces multiple amplification products, each representing a discrete genetic locus, which can be resolved easily by agarose gel electrophoresis. RAPD-PCR has shown promise for use in a wide variety of organisms including bacteria, higher plants, vertebrates, and invertebrates including mosquitoes and other insects. It has been used as a tool for genetic mapping; strain, species and population identification and systematics (Hadrys et al. 1992, Chapco et al. 1992, Black et al. 1992, Kambhampati et al. 1992, Williams et al. 1993, Bowditch et al. 1993, Perring et al. 1993, Gawel & Bartlett 1993, Wilkerson et al. 1993, Wilkerson et al. 1995). Materials and Methods Identification - Peyton et al. (1992) provide a definition of the subgenus Nyssorhynchus. Species in the Albitarsis Complex have hindtarsomere 5 without a basal dark band and vein 1A mostly pale-scaled (Argyritarsis Section) and sternum I with a distinct row of white scales on each side with posterolateral scale tufts beginning on segments III or IV. The sister species An. braziliensis (Chagas) has tufts beginning on segment II [see Linthicum (1988) for additional immature characters]. Specimen preservation - Adults and larvae used in DNA analysis were initially frozen in liquid nitrogen and transported to our laboratory on dry ice where they were maintained at -70 C. Larval and pupal exuviae were collected from individually reared adults and preserved in 80% ethanol and held for slide mounting. Each individually reared specimen was given a unique code number that associated it with other progeny of a single female and also associated it with a pin-mounted adult. Source of specimens - Collection localities of F1 progeny broods used in this study are given in Table I along with those presented in Wilkerson et al. (1995). Associated specimens retained for morphogical study are deposited in the National Museum of Natural History, Smithsonian Institution, Washington DC; US Army Medical Research Unit, Rio de Janeiro; Instituto Oswaldo Cruz, Rio de Janeiro; and the Núcleo de Pesquisa Taxonômica e Sistembetatica em Entomologia Modica (NUPTEM) at the Universidade de Spio Paulo. Experimental procedures - DNA Isolation. Individual larvae or adults were ground with a plastic pestle in microcentrifuge tubes in 200 ul extraction buffer (100mM Tris pH 8.0, 100mM EDTA, 100mM NaCl); proteinase K was then added to 200 ug/ml and SDS to 0.5%. After incubation at 55 C for 3 to 12 hr, RNase was added to a final concentration of 100 ug/ml and incubated at room temperature for 30 min. The solution was extracted once with an equal volume of phenol/chloroform/ isoamyl alcohol (25:24:1, equilibrated with 10 mM Tris pH 8.0, 1mM EDTA) by heating to 55 C for 10 min with periodic mixing of phases. After brief centrifugation in a microcentrifuge, the supernatant was extracted with chloroform/isoamyl alcohol (24:1) as above. The supernatant was collected, 2 volumes (400ul) 95% ethanol were added to it, and the solution stored at - 20 C for 15 min to precipitate the DNA. The DNA was pelleted (15,900 x g in a microcentrifuge for 4 min), washed with 700 ul 70% ethanol, dried under vacuum, and dissolved in 100 ul 10 mM Tris pH 7.5, 1mM EDTA. Typical yields were 0.5 to 6.5 ug DNA per individual. RAPD PCR amplification. Detailed procedures are discussed in Bowditch et al. (1993). Total reaction volumes of 25 ul were used with the following final concentrations: 11 mM Tris-HCl (pH 8.3); 50 mM KCl; 1.9 mM MgCl2; 0.1 mg/ml BSA; 0.1 mM each of dATP, dCTP, dGTP and TTP; 0.24 pmol/ul (6 pmol) primer; 0.2-4.0 ng/ul template DNA; and 0.02-0.06 U/ul (0.5-1.5 units) Taq DNA polymerase. PCR conditions. A Perkin-Elmer Cetus model 480 thermocycler was used for all reactions with the following parameters: one min denaturation at 94 C followed by 45 cycles of one min at 94 C, annealing one min at 35 C and elongation two min at 72 C, all with minimum ramp times. Agarose gel electrophoresis. Using standard methods (Sambrook et al. 1989), amplification products were analyzed using a USA Scientific "Extra Wide" mini horizontal system (cat. no. 3488-0000) which uses 100 ml 1.5% agarose minigels with 0.8 ug/ml ethidium bromide run at 50 volts/25 milliamps for about 3 hr in TBE (89mM Tris base, 89mM boric acid and 2mM EDTA, pH 8.3). Amplification products were observed and photographed using short wave (312 nm) ultraviolet light. Molecular weight standards were provided by lambda DNA digested with Hind III, and FX174 DNA digested with Hae III (New England Biolabs). The approximate molecular weight of amplification products was calculated using a program written for Lotus 1-2-3 by AF Cockburn, United States Department of Agriculture, Gainesville, Florida, USA. Oligonucleotide primers. All primers were 10 bases in length. Primers were purchased from Operon Technologies, Alameda CA. Primers discussed in the text have the following sequences (5'-3'): B16, TTTGCCCGGA; B05, TGCGCCCTTC; D01, ACCGCGAAGG; C07, GTCCCGACGA; B02, TGATCCCTGG; A01, CAGGCCCTTC; C19, GTTGCCAGCC; C16, CACACTCCAG; C15, GACGGATCAG; B11, GTAGACCCGT; A12, TCGGCGATAG; A08, GTGACGTAGG. Based on the results of Wilkerson et al. (1995) (Figs 2-13) the following primers were used to amplify species-specific markers for identification of unknown specimens: Sp A, primers A01, B16, (2 markers); Sp B, primers C07, B02, C15 (4 markers); Sp C, primers A01, C16, C19, D01 (7 markers); and Sp D, primers A12, D01 (3 markers). In addition, the presence or absence of shared bands produced by primers B05 (species A,C), C19 (species A,B,D) and D01 (species C,D) were also taken into consideration (Table II). In order to make direct comparison with previous results, known specimens were run with all unknowns. To minimize the number of reactions needed for an identification, a "best guess" of a species identity was first made based on published and unpublished analyses of morphological characters [e.g. larval seta 3-C branched in An. (Nys.) deaneorum (Rosa-Freitas 1989) or length of basal dark band on hindtarsomere 2 and the relative length of the prehumeral dark spot (unpublished data)], or based on what is already known about the distributions of the species (e.g. species A is only recorded from southern Brazil, Bolivia, Paraguay and northern Argentina while species B has not been recorded from western Brazil). A species-specific primer was then used for the suspected species. Those matching were verified with one or two additional species-specific primers. Those that didnAEt match were tested with a next "best guess". Figure 14 shows species C primer A12 run with a series of specimens from Venezuela (lanes 4-18) and a single specimen from the State of Bahia, Brazil (lane 3). The known specimen of species C is in lane 2 from the type locality of An. (Nys.) marajoara, Ilha de Marajo, State of Parbeta, Brazil. The specimen in lane 3 that did not match was then run with species B primers with which it did match (not shown). Figure 15 shows a series of specimens from the type locality of An. (Nys.) deaneorum, Guajarbeta-Mirim, State of Rondonia, Brazil (lanes 3-11 and 13) which, when run with species D primer A12, compared positively with a known species D from Guaíra, State of Sao Paulo, Brazil (lane 2). The specimens that did not match (lanes 12 and 14-19) from Peixoto de Azevedo, State of Mato Grosso, Brazil are among those run with species C primer C19 shown in Fig. 16 (lanes 3-9). RESULTS Based on genetic markers determined by Wilkerson et al. (1995) (Figs 2-13; Table II) we were able to identify the four known species belonging to the Albitarsis Complex from an additional 12 sites in Brazil and 4 in Venezuela (Table I; Fig 1). An additional eight individuals of species A, 41 of species B, 108 of species C and 26 of species D were identified. All (except nine species C) represent separate progeny broods with associated larval and pupal skins and pinned adults. To carry out identifications we used ten of the 12 primers used by Wilkerson et al. (1995) (Table II; Figs 2-13). These primers produced correlated markers as follow: A(3), D(3), B(4) and C(7). These data validate previous results (Wilkerson et al. 1995) with some variation (Table III). Although it is preferable to use absolutely "fixed" markers for identifications, in reality this is probably not possible when one uses larger samples and/or widely separated populations. It is not critical for the effective use of RAPD markers, to have 100% fixed markers, but to have nearly fixed markers that are correlated. As long as all the exceptional missing markers do not correspond in a single individual, there should be no confusion about assigning individuals to their respective genetically defined taxa. A comparison of the frequency of diagnostic markers between Wilkerson et al. (1995) and the present study show some differences. The 0.38 kbp band produced by primer C07 and the 1.88 kbp band produced by primer B02 were missing in a small percentage of specimens reported here for the first time. Also, the 1.41, 1.22 and 0.85 kbp bands produced by primer A01 for species C were found in slightly lower frequencies than before. Wilkerson et al. (1995) reported that only one individual out of 61 had two of the three bands missing. In the present study, two of 108 also lacked these same two bands (1.22 and 0.85 kbp). However, the other four species C markers tested for by primers C16, D01 and C19 were present in both individuals. The 1.00 kbp marker produced by primer C15 in species B was the only one found to be absent in a significant proportion of individuals compared to the previous study (100%, 15/15 vs 68.3%, 28/41). In addition, a 1.41 kbp band produced by this primer which was not recognized as a potentially useful species-specific marker because of its inconsistent brightness, was actually consistently present and recognizable in all individuals tested. A possible homologous 1.41 kbp band was seen in four of fifteen species C, but further testing was not done to verify its size. Twenty-three progeny broods from the type locality of An. (Nys.) deaneorum matched for all three markers previously determined for species D. Even though the initial species D sample from northern Argentina and southern Brazil was small (n = 9), these markers were also present in the larger sample (n = 26) from two localities in the State of Rondonia, Brazil. A distance of approximately 2,400 km was previously reported (Wilkerson et al. 1995) between populations of species C. With the addition of specimens of species C from Venezuela, the maximum distance we have found between populations exhibiting the same diagnostic markers is now approximately 4,620 km (Iguape, State of Spio Paulo, Brazil to Rio Socuavo, State of Zulia, Venezuela). Species C and D were sympatric at Costa Marques, State of Rondônia, Brazil. These two species had been collected previously in the same human and bovine bait collections (Klein & Lima 1990). With this finding, all possible pairs of species, except species A and C, have been found to be sympatric at some locality. The site near Guaíra, State of Paranbeta, Brazil remains the only site sampled with three of the species occurring together (A, B, D). DISCUSSION Using RAPD-PCR we have begun to solve parts of the Albitarsis Complex puzzle. However, the relation of most previous epidemiological, biological and morphological studies to our results cannot be determined with certainty (also discussed in Wilkerson et al. 1995). Many have attempted to correlate malaria transmission with behavior and/or relative length of the dark portion of hindtarsomere 2 (reviewed by Rosa-Freitas & Deane 1989 and Rosa-Freitas et al. 1990). Some of the studies demonstrated a correlation, while most were ambiguous. These attempts had their basis in early observations regarding malaria transmission and biting-resting behavior. Galvpio and Damasceno (1944) regarded An. (Nys.) albitarsis albitarsis as exophilic, not related to malaria transmission and to have 77-84% of hindtarsomere 2 dark. The only specimens we have seen that agree with this description of hindtarsomere 2 are described here as species A, "true" An. (Nys.) albitarsis. At the same time, Galvpio and Damasceno (1944) described as new An. (Nys.) albitarsis domesticus, regarding it as endophilic, related to malaria transmission and to have 36-50% of hindtarsomere 2 dark. As a basis for their description they contrasted domesticus to An. (Nys.) albitarsis albitarsis. An. (Nys.) albitarsis domesticus was later synonymized under An. (Nys.) marajoara by Linthicum (1988). Since the type locality for both species is the same, and because no type specimens apparently exist for An. (Nys.) albitarsis domesticus to refute this, we agree with Linthicum AE's synonymy (previously reported in Wilkerson et al. 1995). Even though there is a temptation to use such a useful descriptive name as "domesticus" for members of this complex that enter houses and transmit malaria, it should no longer be used since there may be several species which fit this description, to which none the name "domes-ticus" can be applied. After sampling 18 widely separated populations in Brazil, Rios et al. (1984) also came to the conclusion that it was impossible to separate the two putative subspecies, An. (Nys.) albitarsis albitarsis and An. (Nys.) a. domesticus, using these same "traditional" characters. Anopheles (Nys.) marajoara sensu Linthicum (1988) is based primarily on specimens from Panama, not the type locality, Ilha de Marajo, Brazil. He included all specimens that he studied, except An. (Nys.) albitarsis s.s., ranging from Costa Rica to southern Brazil, under the name An. (Nys.) marajoara, including those that were later described as An. (Nys.) deanorum. His discussion points out the great variation he found which strongly suggests that he was dealing with a mixture of taxa. Previous cytogenetic and molecular studies have offered some insight into understanding this complex. It is not clear how these findings relate to those presented here because in most cases voucher specimens are not available for comparison. Kreutzer et al. (1976) found three chromosomally differentiated populations, "C" in Colombia and Venezuela and two others, B1 and B2, ranging from Rio de Janeiro to Belom and just north of Ilha de Marajo. We do not believe that we have yet sampled their population "C". Steiner et al. (1982), using isozyme analysis, found three "genetically differentiated" populations at sites from Sao Paulo to just west of Marajo Island based on 12 allozyme loci and chromosome analysis. According to their map, species "a" and "b" were found near the coast and "b" was also found inland but, according to their Table II, "a" was also found with "b" inland at Logoa (sic, Lagoa?) Feia. An additional discrepancy failed to show species "b" at Araraquara, sympatric with their species "c". These possibly correspond to "B1" and "B2" of Steiner et al. (1982). However, we were not able to locate published information on chromosomes cited in their analysis. In a study of 10 populations Rosa-Freitas et al. (1990) found no morphological differences, except for An. (Nys.) deaneorum, but through isoenzyme analysis concluded that there were five "groups" in Brazil and Argentina. Narang et al. (1993) compared four populations in the Albitarsis Complex in Brazil using 18 allozyme loci and mitochondrial DNA restriction fragment length polymorphisms. They reported that two morphologically distinct taxa existed, An. (Nys.) deaneorum and An. (Nys.) marajoara, the latter made up of 2 or 3 biochemically recognizable forms. One of the forms of An. (Nys.) marajoara was sympatric with An. (Nys.) deaneorum at Costa Marques, State of Rondonia, and the other one or two forms of An. (Nys.) marajoara, depending on interpretation, were from Iguape, State of Spio Paulo, and from its type locality of Marajo Island, State of Parbeta. The Iguape specimens are the same as those used by us in the present study (species C) and may represent a species responsible for a malaria outbreak in Iguape reported by Schiavi (1945). As reported here the two species from Costa Marques (C, D) are probably An. (Nys.) marajoara and An. (Nys.) deaneaorum. RAPD-PCR has proven to be effective in separating cryptic Anopheles species and for identifying unknown specimens. It has the potential for use as a routine diagnostic tool for identification of Anopheles sibling species, but because of the sensitivity of RAPD-PCR to changing experimental conditions, care should be taken in its use (Williams et al. 1993, Black 1993). It has the advantage of relatively low initial investment of time for determining species-specific markers when compared to designing and testing custom-made species-specific PCR primers based on known nucleotide sequences.
TABLE I
Summary of collection localires for Albitarsis Complex species
identified by Random Amplified Polymorphic DNA -
Polymerase Chain Reaction
State/ Locality Coordinates
Date
Province
--------------------------------------------------------------
BRAZIL
Rondonia^a Costa Marques 12 28'S 64 16'W
28-III-92
Rondonia^a Guajara-Mirim 10 50'S 65 20'W
26-III-92
Para^a Belem 1 27'S 48 29'W
8-X-92
Para^a Primavera 0 56'S 47 06'W
16-X-92
Mato Grosso^a Matupa, Peixoto 10 23'S 54 54'W
20-IV-93
de Azevedo
Bahia^a Itaquara 13 26'S 39 56'W
30-I-93
Ceara^a Fortaleza 3 43,S 38'30'W
8-III-93
Ceara^a Parapaiba a 3 25,S 39 13'W
9-III-93
Santa Catarina^ Massaranduba a 26 35'S 48 58'W
8-I-93
Rio de Janeiro^ Morro da Panela 22 58,S 43 21'W
19-193
Arnazoilas^a Manaus 2 53.S 60 15'W
16-XII-93
Sao Paulo Ilha Comprida 24 42.8'S 47 31.6'W
9-II-89
Sao Paulo 6 km SW Registro 24 36.8'S 47 53.1'W
26-I-92
Sao Paulo Ponte Melo Peixoto 22 39.05,S 53 01'W
10-II-92
Para Capanema 1 24,S 47 11'W
VII-93
Para Ilha de Marajo l OffS 49 30'W
VIII-91
Parana Santa Helena 24 56'S 54 23'W
30-I-92
Parana nr. Guaira 24 04'S 54^ 15'W
1-II-92
Espirito Santo Aguia Branca 18 59'S 40 44'W
20-I-92
VENEZUELA
Zulia^a Rio Socuavo 8 54'N 72 38'W
22-IX-92
Zulia^a Tres Bocas 78 54'N 72 38'W
24-IX-92
Cojedes^a Finca Rosa Blanca 9.6 N 68.9 W
26-VI-91
Barinas^a Castilia 8 25~q 70 38'W
14-IX-92
PARAGUAY
Alto Parana Rio Acaray 25 29,S 54 42'W 4-II-92
Alto Parana Hernandarias 25 22,S 54 45'W 6-II-92
Alto Parana nr. National (not known) 8-11-92
ARGENTINA
Misiones Posadas 27 23,S 55 53'W
30-I-92
Corrientes 90km W Posadas (not known)
31-I-92
Corrientes Corrientes 27 28,S 59 50'W
31-I-92
Corrientes 11 km W Mercedes 29 12,S 58 05'W
1-II-92
Corrientes Santa Tome 28 33,S 56 03'W
2-II-92
Buenos Aires Baradero 33 48,S 59 30'W
6-II-92
TABLE 1 contd./
State/Province Species(n) Map Ref.
----------------------------------------------
BRAZIL
Rondonia^a C(16), D(3) 13
Rondonia^a D(23) 12
Para^a B(2) 8
Para^a B(5) 9
Mato Grosso^a C(29), B(12) 14
Bahia^a B(9) 15
Ceara^a B(11) 1
Ceara^a B(13) 10
Santa Catarina^ A(8) 26
Rio de Janeiro^ B(1) 18
Arnazoilas^a C(34) 7
Sao Paulo C(18) 24
Sao Paulo A(30), B(19) 25
Sao Paulo A(l) 17
Para c(8), B(1) 6
Para C(43) 5
Parana A(5), D(I) 20
Parana A(12), B(I), D(4) 19
Espirito Santo B(I) 16
VENEZUELA
Zulia^a C(16) 1
Zulia^a C(I) 2
Cojedes^a C(I0) 4
Barinas^a C(2) 3
PARAGUAY
Alto Parana A(9) 22
Alto Parana A(8), B(6) 21
Alto Parana B(9) 2
ARGENTINA
Misiones A(12) 27
Corrientes A(1), D(2) 28
Corrientes A(9), D(2) 29
Corrientes A(5) 31
Corrientes A(8) 30
Buenos Aires A(22) 32
TABLE II
Summary of Randon Amplified Polymorphic DNA
markers used to identify species of the Albitarsis
Complex
----------------------------------------------------
Primer Approximate A B C D
kbp
----------------------------------------------------
B16 1.75 +a - - -
B05 1.74 + - + -
DO1 2.2-2.3 + + + +
0.96 - - - +a
0.80 - - + +
0.55 - - +a -
C07 0.38 - +a - -
B02 2.88 - +a - -
A01 2.03 +a - - -
1.41 - - +a -
2.22 - - +a -
0.85 - - +a -
C19 1.30 + + - +
1.17 - - +a -
0.88 - - +a -
C16 0.99 - - +a -
C15 1.41 - +a - -
1.24 - - - +a
1.00 - +a - -
0.86 +a - - -
B11 1.78 + + + -
1.26 - - - +a
A12 1.29 - - - +a
1.09 - - - +a
A08 1.09 - - - +a
a: species specific markers; +: marker in common to
more than one species; -: marker not present species
TABLE III
Diagnostic primers used in identification of Albitarsis
Complex species and comparison to Wilkerson (1995) results
Species Primer Appox. Wilkerson (1995) Present study
kbp
-------------------------------------------------------------
A B05 1.74 121/121 (100%) 8/8 (100%)
B16 1.75 81/82 (98.8%) 8/8 (100%)
A01 2.03 118/118 (100%) 8/8 (100%)
B C07 0.38 32/32 (100%) 40/41 (97.5%)
B02 1.88 35/35 (100%) 38/41 (92.6%)
C15 1.00 15/15 (100%) 28/41 (68.3%)
1.41 15/15 (100%) 41/41 (100%)
C C16 0.99 63/63 (100%) 66/66 (100%)
D01 0.55 42/42 (100%) 29/29 (100%)
A01 1.41 59/61 (96.7%) 91/108 (84.3%)
1.22 58/61 (95.1%) 86/108 (79.6%)
0.85 59/61 (96.7%) 100/108 (92.3%)
C19 1.17 58/58 (100%) 84/84 (100%)
0.88 58/58 (100%) 84/84 (100%)
D
A12 1.29 9/9 (100%) 26/26 (100%)
1.09 9/9 (100%) 26/26 (100%)
D01 0 96 9/9 (100%) 26/26 (100%)
Figure 1
Figure 2
Figure 3
Figure 4
Figure 5
Figure 6
Figure 7
Figure 8
Figure 9
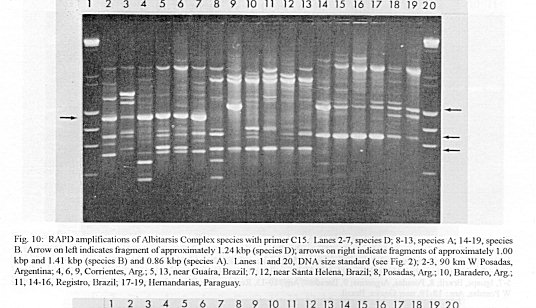
Figure 10
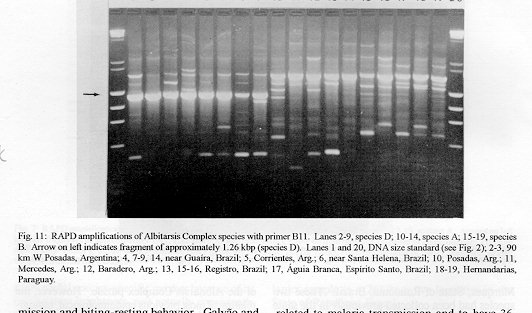
Figure 11
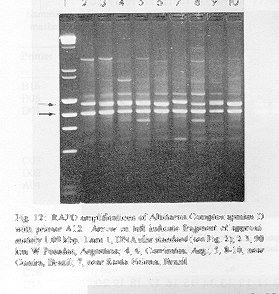
Figure 12
Figure 13
Figure 14
Figure 15
Figure 16
ACKNOWLEDGMENTS To the personnel of the U.S. Army Medical Research Unit, Rio de Janeiro, Brazil, for collecting, rearing and curating many of the specimens used in this study. They are: Eric G Milstrey, Maria de Fbetatima Barros, Joso Rodrigues Vargas and Temizio Rodrigues Pereia. Also in Brazil, Agostinho Marques Cruz, Fundacaio Nacional da Saude, Brasilia, provided invaluable administrative support as did Jorge Arias, Pan American Health Organization, Brasília. To Soma Mitra for her conscientious and careful DNA analyses. To Jan Conn and Phil Lounibos (supported by NIH grant AI-31034) who kindly provided specimens from Venezuela. REFERENCES Black WC III 1993. PCR with arbitrary primers: approach with care. Insect Mol Biol 2: 1-6. Black WC III, Duteau NM, Puterka GJ, Nechols JR, Pettorini JN 1992. Use of the random amplified polymorphic DNA polymerase chain reaction (RAPD-PCR) to detect DNA polymorphisms in aphids (Homoptera: Aphididae). Bull Ent Res 82: 151-159. Bowditch BM, Albright DG, Williams JGK, Braun MJ 1993. The use of RAPD markers in comparative genome studies. Meth Enzymol 224: 294-309. Chapco W, Ashton NW, Martel RKB, Antonishyn N 1992. A feasibility study of the use of random amplified polymorphic DNA in the population genetics and systematics of grasshoppers. Genome 35: 569-574. Galvpio ALA, Damesceno RG 1942. Sobre um novo anofelino da Ilha de Marajó, Anopheles (Nyssorhynchus) marajoara. Folia Clin Biol, Spio Paulo 14: 60-66. Galvpio ALA, Damesceno RG 1944. Observacoes sobre anofelinos do complexo albitarsis (Diptera Culicidae). Ann Fac Med, Spio Paulo 20: 73-87. Gawel NJ, Bartlett AC 1993. Characterization of differences between whiteflies using RAPD-PCR. Insect Mol Biol 2: 33-38. Hadrys H, Balick M, Schierwater B 1992. Applications of random amplified polymorphic DNA (RAPD) in molecular ecology. Mol Ecol 1: 55-63. Kambhampati S, Black WC IV, Rai KS 1992. Random amplified polymorphic DNA of mosquito species and populations (Diptera: Culicidae): Techniques, statistical analysis, and applications. J Med Entomol 29: 939-945. Klein TA, Lima JB 1990. Seasonal distribution and biting patterns of Anopheles mosquitoes in Costa Marques, Rondônia, Brazil. J Amer Mosq Cont Assoc 6: 700-707. Kreutzer RD, Kitzmiller JB, Rabbani MG 1976. Cytogenetically distinguishable sympatric and allopatric populations of the mosquito Anopheles albitarsis. Acta Amaz 6: 473-481. Linthicum KJ 1988. A revision of the Argyritarsis Section of the subgenus Nyssorhynchus of Anopheles (Diptera: Culicidae). Mosq Syst 20: 98-271. Lynch-Arribalphalzaga F 1878. Descripcion de tres nuevos Culicidae de Buenos Aires. Nat Argent 1: 149-152. Narang SK, Klein TA, Perera OP, Lima JB, Tang AT 1993. Genetic evidence for the existence of cryptic species in the Anopheles albitarsis complex in Brazil: Allozymes and mitochondrial DNA restriction fragment length polymorphisms. Biochem Gen 31: 97-112. Perring TM, Cooper AD, Rodriguez RJ, Farrar CA, Bellows TS 1993. Identification of a whitefly species by genomic and behavioral studies. Science 259: 74-77. Peyton EL, Wilkerson RC, Harbach RE 1992. Comparative analysis of the subgenera Kerteszia and Nyssorhynchus of Anopheles (Diptera: Culicidae). Mosq Syst 24: 51-69. Rios RI, Nascimento LZ, de Oliveira AC 1984. Complexo Anopheles (Nyssorhynchus) albitarsis: impossibilidade de separbeta-lo em duas subespocies, a. albitarsis albitarsis e A. albitarsis domesticus (Diptera Culicidae). Rev Brasil Bio 44: 461-465. Rosa-Freitas MG 1989. Anopheles (Nyssorhynchus) deaneorum: A new species in the albitarsis complex (Diptera: Culicidae). Mem Inst Oswaldo Cruz 84: 535-543. Rosa-Freitas MG, Deane LM 1989. The neotype of Anopheles albitarsis (Diptera: Culicidae). Mem Inst Oswaldo Cruz 84: 289-302. Rosa-Freitas MG, Deane LM, Momen H 1990. A morphological, isoenzymatic and behavioural study of ten populations of Anopeles (Nyssorhynchus) albitarsis Lynch-Arribalzaga, 1878 (Diptera: Culicidae) including from the type-locality - Baradero, Argentina. Mem Inst Oswaldo Cruz 85: 275-289. Schiavi A 1945. Nota sobre mosquitos vetores em Iguape. Arq Hig Saud Publ, Spio Paulo 10:69-75. Sambrook J, Fritsch EF, Maniatis T 1989. Molecular Cloning. A laboratory manual, 2nd edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. Steiner WWM, Narang S, Kitzmiller JB, Swofford DL 1982. Genetic diversity and evolution in neotropical Anopheles (subgenus Nyssorhynchus), p. 523-550, In WWM Steiner et al. (eds.), Recent Developments in the Genetics of Insect Disease Vectors. Stipes, Champaign, IL. Welsch J, McClelland M 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucl Acids Res 18: 7213-7218. Wilkerson RC, Parsons TJ, Albright DG, Klein TA, Braun MJ 1993. Random amplified polymorphic DNA (RAPD) markers readily distinguish cryptic mosquito species (Diptera: Culicidae: Anopheles). Ins Mol Biol 1: 205-211. Wilkerson RC, Parsons TJ, Klein TA, Gaffigan TV 1995. Discovery by RAPD-PCR of cryptic species belonging to the Anopheles (Nyssorhynchus) albitarsis complex from Paraguay, Argentina and Brazil (Diptera: Culicidae). J Med Entomol (in press). Williams JGK, Kubelik AR, Livak KJ, Rafalski JA, Tingey SV 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nuc Acids Res 18: 6531-6535. Williams JGK, Hanafey MK, Rafalski JA, Tingey SV 1993. Genetic analysis using random amplified polymorphic DNA markers. Meth Enzymol 218: 704-740. The views of the authors do not purport to reflect the views of the supporting agencies. +Corresponding author. Present address: Museum Support Center, Smithsonian Institution, Washington DC 20560. Received 2 January 1995 Accepted 8 August 1995 Copyright 1995 Fundacao Oswaldo Cruz, FIOCRUZ
The following images related to this document are available:Photo images[oc95147g.jpg] [oc95147i.jpg] [oc95147h.jpg] [oc95147e.jpg] [oc95147j.jpg] [oc95147k.jpg] [oc95147l.jpg] [oc95147o.jpg] [oc95147c.jpg] [oc95147p.jpg] [oc95147b.jpg] [oc95147n.jpg] [oc95147d.jpg] [oc95147m.jpg] [oc95147f.jpg]Line drawing images[oc95147a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}