 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz Vol. 90(6), Nov./Dec. 1995 Lignanes from the Brazilian Melia azedarach, and their Activity in Rhodnius prolixus (Hemiptera, Reduviidae) Marise MO Cabral, Eloi S Garcia, Alphonse Kelecom* Departmento de Bioquimica e Biologia Molecular, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil *Departamento de Biologia Geral, Universidade Federal Fluminense, Caixa Postal 100.436, 24001-970 Niteroi, RJ, Brasil
Code Number: OC95152
Size of Files:
Text:
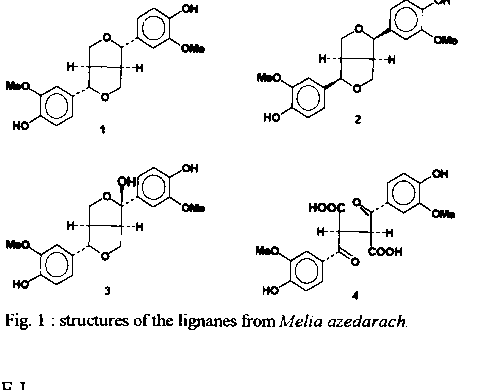
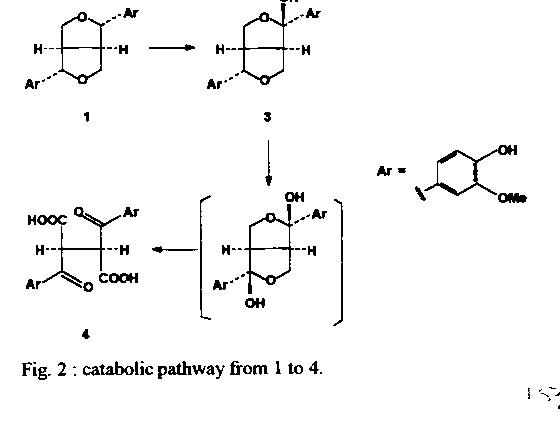
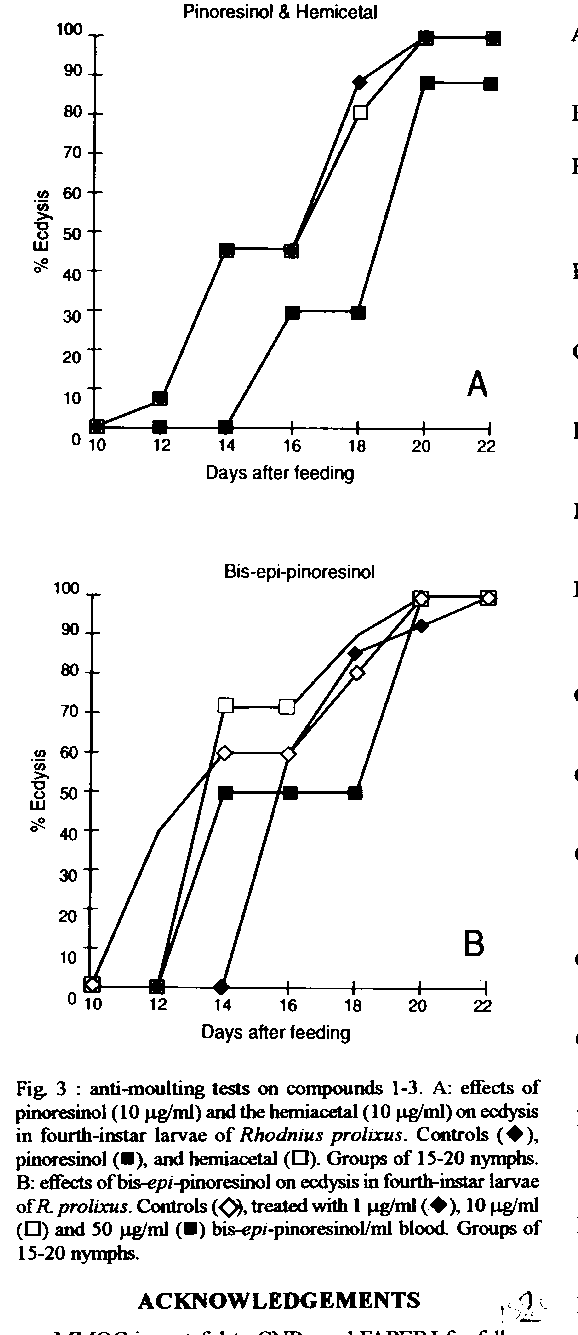
Graphics: Line Drawings (gif) - 27KA study of the phagoinhibitor and anti-moulting activities of the Brazilian Melia azedarach, collected in the State of Rio de Janeiro, Brazil, allowed the isolation of four lignanes identified as pinoresinol (1), bis-epi-pinoresinol (2), the hemicetal (3) and the diacid (4). These substances are devoid of anti-moulting activity. Key words : lignanes - Meliaceae - Melia azedarach - insects - Rhodnius prolixus Chagas disease is widely spread in South America. Dissemination of the illness involves haematophagous insects, such as Rhodnius prolixus and some other related ones. It is long known that human contamination occurs through the feces that triatomid infested insects invariably deposit on feeding (Garcia & Azambuja 1991). Obviously, control of the disease depends heavily upon the efficient control of its vectors. A number of insecticides are commonly used for that purpose. They offer a series of disadvantages since they are usually both highly toxic and non-specific. In addition, most of them are not biodegradable and must thus be considered as polluting agents (van Beek & de Groot 1986). Safer alternatives may be found among natural sources, mainly higher plants that afford a number of repellent and toxic secondary metabolites (Subrahmanyam 1990). Abundant literature describes plants of the family Meliaceae that produce meliacins (or limonoids), some of which are endowed with potent phagorepellent properties (Nakanishi 1976). For example, azadirachtin-A, a highly oxygenated tetra-nor-triterpenoid, has been isolated from the Indian Azadirachta indica (Butterworth & Morgan 1968) and from the Kenyan Melia azedarach (Morgan & Thornton 1973). This compound is a remakable phagoinhibitor (40 mg/ml) with anti-moulting (DE50 4.10-4 mg/ml) activities on R. prolixus (Garcia et al. 1984), and has turned out to be a potential useful drug for Chagas disease control. However, production of azadirachtins might suffer from strong biogeographic dependance since it has been reported recently that crude extracts of M. azedarach from Paraguay were devoid of any anti-moulting activity (Arias & Hirschmann 1988). Only fatty acids, some classical phytosterols and two coumarins, scoparone and isofraxidin, could be isolated (Arias & Hirschmann 1988). Thus, it seemed of great interest to start a study of the Brazilian M. azedarach, in order to look for the presence of anti-moulting principles. This exotic plant, of African origin, is widely cultivated in Brazil where it is known as “cinamomo”. As far as we are aware, there seems to be no report on the isolation of anti-ecdysis compounds from the Brazilian plant. This paper reports on the structure identification and bioassays of four lignanes isolated from a bioactive fraction of cinamomo seeds. MATERIALS AND METHODS Extraction and fractionation - Seeds of M. azedarach (1,300 g), collected in the neighbour-hood of Niteroi (State of Rio de Janeiro, Brazil), were extracted exhaustively with MeOH (2 x 5 l). Filtration and evaporation of the solvent under reduced pressure furnished a gummy residue (245g). Partition between hexane and 5% aq.MeOH, followed by evaporation of the methanolic phase and treatment with EtOAc afforded an active fraction "A" (71 g). Bioassay guided fractionation of a 1g aliquot of "A", by conventional chromatographic processes using silica gel and mixtures of CHCl3-MeOH, furnished the active fraction, F-58 (21 mg) that contained phytosterols, four lignanes (1-4) and one triterpene whose structure elucidation will be reported elsewhere. Preparative scale purification of "A" was carried out as follows. Fraction "A" (49 g) was coated on silica gel, placed in a Buchner filter and submitted, under reduced pressure, to selective desorption using successively hexane, CHCl3 and acetone. Fractions 1-3 were combined, affording "B" (11 g) that was submitted to quick filtration on silica gel (eluent : step gradient of acetone 2-5-10-20-50-80 and 100% in CHCl3). Fractions 6-7 ("M", 2 g) from the latter filtration contained lignanes 1 and 2, phytosterols and one triterpene. Fractions 9-11 ("E", 300 mg) contained the more polar lignanes 3 and 4. Fraction “M” was submitted to conventional silica gel column chromatography (eluent : gradient of AcOEt in hexane from 2 to 50%). Lignanes 1 (50 mg) and 2 (30 mg) were obtained both better than 90% pure. Finally, fraction "E" was submitted to preparative HPLC purification on a RP-8 column (theta 50mm), eluted with MeOH-H2O 45:55, at a pressure of 32 bars, under continuous flow of 100ml/min and UV detection at 215nm. This allowed the isolation of lignane 4 (fractions E1-E2, 10.5 mg, 91% pure) and lignane 3 (fraction E8, 4.5 mg, 98% pure). Bioassays - Fourth instar nymphs of R. prolixus were used. Test material was dissolved in EtOH-saline (1:4). Aliquots were added to blood in order to obtain desired final concentrations (10 to 100 mg/ml). Test blood was placed in special designed feeders (Garcia & Rembold 1984), and the insects were allowed to feed. After feeding, the insects were weighted, incubated at 28 C and observed every two days over a 1 month period. Only fully fed insects were used ; partially fed ones were discarded. Death and ecdysis were counted. RESULTS We observed both phagoinhibitor and anti-moulting activities in the methanolic crude extract of the seeds of M. azedarach. Bioassay-guided fractionation of this crude extract, by solvent-solvent partition, furnished an EtOAc phase that contained almost all the activity. Repetitive silica gel column chromatographies yielded the active fraction F-58 that inhibited 100% moulting of R. prolixus at 25 mg/ml blood (Cabral et al. in preparation). HPLC analysis, on a RP-18 column, indicated the presence, in the crude extract, of traces of a constituent whose retention time was identical to an authentic sample of azadirachtin-A (Cabral et al. in preparation). Purification of this active fraction by reversed phase HPLC yielded classical phytosterols (cholesterol, campesterol and stigmasterol), one triterpene and four lignanes (1-4, Fig. 1). Compound 1 was unequivocally identified as the well known pinoresinol, on the basis of its 1H and 13C NMR spectra (Tables I, II) identical with literature data (Fonseca et al. 1979, Marcos et al. 1990 , Cuenca & Catalan 1991), and also from direct comparison with the ^1H and ^13C NMR spectra of an authentic sample. Compound 2 had a structure very close to that of pinoresinol (1), showing almost the same 1H NMR spectrum (Table I) with only slight differences in the pattern of the aromatic protons. Since epimerization at one of the benzylic positions should change the pattern of H-7,7' from a 2H doublet at delta 4.71 into two 1H doublets at d 4.71 and 4.55 (Pelter et al. 1976, Marcos et al. 1990), and since the ^1H NMR spectrum showed that 2 maintained the symmetry of 1, we propose for 2 the structure bis-epi-pinoresinol. This structure needs further confirmation. Compound 3 again showed, in ^1H NMR, the same aromatic moiety as 1 and 2, and the presence of signals corresponding to the bis-perhydrofurane system. However, integration of the benzylic signal indicated the presence of only one such proton (H-7); carbon-7' is thus quaternary. We propose for this metabolite the hemicetal structure 3 derived from pinoresinol. This hypothesis, although reasonable, could not be corroborated by 13C NMR spectroscopy due to the very low amount of available pure sample and must be considered as provisional. Finally, the highly polar compound 4 again showed, in 1H NMR, a symmetrical structure having a tri-substituted aromatic ring and an OMe group. However, major differences were observed between the ^1H NMR spectra of 4 and 1-3. Thus, the low field positions of H-2,2' and H-6,6' indicated the presence of a deshielded group at the C-7,7' positions, probably a carbonyl function. This hypothesis was in agreement with the absence of the 2H doublet attributable to the H-7,7'Æ signals. In addition, the 4H AB system of H-9,9'eq and H-9,9'ax is also absent and the H-8,8' signal, a multiplet at d 3.10 in 1, appears now as a sharp singlet at d 3.65. All this pointed to structure 4, that may be either an isolation artifact or a metabolite obtained by oxidative degradation as depicted in Fig. 2. These results are remarkable since, although lignanes were known from the order Rutales, this is the first report of this class of metabolites in the family Meliaceae, and also in the very well studied genus Melia (Gottlieb & Yoshida 1989). Abundant literature reports on the biological activities of lignanes. They were found to be antineoplastic (Brewer et al. 1979, Dewick & Jacson 1981), antimitotic (Gensler et al. 1977), antibacterial (Nakatani et al. 1988), and anti-hypertensive agents (Sih et al. 1976). They also inhibit the enzyme c-AMP phosphodiesterase (Mac Rae & Towers 1984). On insects, lignanes act as antifeedant (Harmatha & Nawrot 1984) and as larval growth inhibitors (Mac Rae & Towers 1984). In addition, juvenile hormone activity has been reported (Bowers 1968).On the other hand, a number of lignanes act synergistically with a variety of insecticides, increasing their toxicity (Haller et al. 1941, 1942, Fales et al. 1970, Matsui & Munakata 1975, Mac Rae & Towers 1984). Despite this broad spectrum of activities, our bioassays (Fig. 3a,b) showed that lignanes 1-3 are not responsible, by themselves, for the anti-moulting activity observed in the crude extact (Cabral et al. in preparation). Synergistic effects are however not excluded and are now under study in our laboratories. In the absence of botanical confusion of the Paraguayan plant commented above (Arias & Hirschmann 1988), our results suggest that production by M. azedarach of anti-moulting principles and other secondary metabolites may be very latitude- dependent.
TABLE I
IH NMR data for compounds 1-4
H 1 1^a 2
--------------------------------------------------------------
H-2,2' 6.87 d 1.5 6.89 d 6.86 d 1.8
H-5,5' 6.88 d 8.5 6.88 d 8.0 6.82 d 8.4
H-6,6' 6.79 dd 8.5; 1.5 6.82 dd 6.75 dd 8.4; 1.8
H-7,7' 4.71 d 4.6 4.73 d 4.3 4.69 d 4.7
H-8,8' 3.08 m 3.10 rn 3.05 m
H-9,9' eq 4.22 dd 6.9; 9.0 4.27 dd 6.9; 9.1 4.19 dd 7.0; 9.3
H-9,9' ax 3.84 dd 3.9; 9.0 3.87 dd 3.8; 9.2 3.85 dd 3.7; 9.3
OMe 3.89 s 3.91 s 3.81 s
OH 5.58 m 5.89 m n.o
Table I contd./
H 3 4
------------------------------------------------------
H-2,2' 6.94 d 1.6 7.57 bs
H-5,5' 6.76 d 8.1 6.80 d 8.5
H-6,6' 6.81 dd 8.1; 1.6 7.52 bd 8.5
H-7,7' 4.70 d 4.4
H-8,8' 3.14 m (?) 3.65 s
H-9,9' eq 4.23 dd 6.9; 9.0 -
H-9,9' ax 3.84 dd 3.6; 9.0 -
OMe 3.85 s 3.88 s
OH n.o. n.o.
a: Cuenca & Catalan (1991); n.o.= not observed; 1 and 2 in
CDCI^3 at 200 MHz; 3 and 4 in CD3OD at 250 MHz.
TABLE II
^13C NMR data of l (CDCl3) compared with
literature data (Fonseca et al. 1979)
C 1 pinoresinol
--------------------------------------
C-l-l' 132.85 s 132.0
C-2,2' 118.93 d 118.5
C-3,3' 114.23 d 114.4
C-4,4' 145.18 s 145.2
C-5,5' 146.66 s 146.8
C-6,6' 108.57 d 108.8
C-7,7' 85.86 d 85.7
C-8,8' 54.11 d 53.7
C-9,9' 71.63 t 71.3
OMe 55.93 s 55.6 ACKNOWLEDGEMENTS MMOC is grateful to CNPq and FAPERJ for fellowships and to Dr OR Gottlieb for fruitful discussions. AK thanks Dr H Rembold (Max-Planck Institut fur Biochemie, Munchen, Germany) for technical support, Dr R Riguera (Dept. of Chemistry, University of Zaragosa, Spain) for original NMR spectra of pinoresinol and related lignanes, Dr R Ottinger (CIREM, Free University of Brussels, Belgium) and Dr Aurelio B Buarque Ferreira (Dept. of Chemistry, Universidade Federal Rural do Rio de Janeiro, Brazil) for the NMR spectra.
REFERENCES
Arias AR, Hirschman GS 1988. The effects of Melia azedarach on Triatoma infestans bug. Fitoterapia 59: 148-149. Bowers WS 1968. Juvenile hormone activity of natural and synthetic synergists. Science 161: 895-897. Brewer FC, Loike JD, Horwitz SB 1979. Conformational analysis of podo-phyllotoxin and its congeners. Structure-activity relationship in microtubule assembly. J Med Chem 22: 215-221. Butterworth JH, Morgan ED 1968. Isolation of a substance that supresses feeding in locust. J Chem Soc Chem Commun: 23-24. Cuenca MR, Catalan CAN 1991. Monoterpenes and lignanes from Mikania saltensis. J Nat Prod 54: 1162-1164. Dewick PM, Jacson DE 1981. Cytotoxic lignanes from Podophyllum and the nomenclature of aryltetralin lignanes. Phytochemistry 20: 2277-2280. Fales JH, Bodenstein OF, Bowers WS 1970. Seven juvenile hormone analogs as synergists for pyrethrins against house flies. J Econ Entomol 63: 1379-1380. Fonseca SF, Nielsen LT, Rúveda EA 1979. Lignanes of Araucaria augustifolia and 13C NMR analysis of some phenyltetralin lignanes. Phytochemistry 18: 1703-1708. Garcia ES, Azambuja P 1991. Development and interactions of Trypanosoma cruzi within the insect vector. Parasitol Today 7: 240-244. Garcia ES, Rembold H 1984. Effects of azadirachtin on ecdysis of Rhodnius prolixus. J Insect Physiol 30: 939-941. Garcia ES, Azambuja P, Forster H, Rembold H 1984. Feeding and molt inhibition by azadirachtins A, B and 7-acetyl-azadirachtin. Z Naturforsch 39c: 1155-1158. Gensler WJ, Murthy CD, Trammell MH 1977. Non-enolizable podophyllo-toxin derivatives. J Med Chem 20: 635-644. Gottlieb OR, Yoshida M 1989. Lignanes, p. 439-511. In JW Rowe Natural Products in Woddy Plants I. Acad. Press, New York. Haller HL, La Forge FB, Sullivan WN 1942. Effect of sesamin and related compounds on the insecticidal action of pyrethrum on house flies. J Econ Entomol 35: 247-248. Haller HL, Mc Govran ER, Goodhue LD, Sullivan WN 1941. The synergistic action of sesamin with pyrethrum insecticides. J Org Chem 7: 183-184. Harmatha J, Nawrot J 1984. Comparison of the feeding deterrent activity of some sesquiterpenes lactones and a lignane lactone towards selected insect storage pests. Biochem Syst Ecol 12: 95-98. Mac Rae WD, Towers GHN 1984. Biological activities of lignanes. Phytochemistry 23: 1207-1220. Marcos M, Jimenez C, Villaverde MC, Riguera R, Castedo L, Stermitz F 1990. Lignanes and other constituents from South and Central American Zanthoxylum species. Planta Medica 56: 89-91. Matsui K, Munakata K 1975. The structure of piperenone, a new insect antifeeding substance from Piper futokadzura. Tetr Letters: 1905-1908. Morgan ED, Thornton MD 1973. Azadirachtin in the fruit of Melia azedarach. Phytochemistry 12: 391-392. Nakanishi K 1976. Insect growth regulators from plants. In Produits Naturels et la Protection des Plantes 41: 185-198. Nakatani N, Ikeda K, Kikuzaki H, Kido M, Yamaguichi Y 1988. Diaryl-dimethylbutane lignanes from Myristica argentea and their antimicrobial action against Streptococcus mutans. Phytochemistry 27: 3127-3129. Pelter A, Ward RS, Rao EV, Sastry KV 1976. Revised structure for epiaschantin and epimagnolin. Tetr Letters: 2783-2788. Sih CJ, Ravikumar PR, Huang Fu-Chih, Buckner C, Whithock Jr H 1976. Isolation and synthesis of pinoresinol diglucoside, a major antihypertensive principle of Tu-Chung. J Am Chem Soc 98: 5412-5413. Subrahmanyam B 1990. Azadirachtin. A naturally occurring insect growth regulator. Proceed Indian Acad Sci 99: 277-288. van Beek TA, de Groot AC 1986. Terpenoids antifeedants I. An overview of terpenoid anti-feedants of natural origin. Rec Trav Chim Pays-Bas 105: 513-527. This work was supported by CNPq, FINEP, CAPES, DAAD and Volkswagen Fundation
Received 14 March 1995 Accepted 12 July 1995 Copyright 1995 Fundacao Oswaldo Cruz, FIOCRUZ
The following images related to this document are available:Line drawing images[oc95152c.gif] [oc95152b.gif] [oc95152a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}