 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz Vol. 90(6), Nov./Dec. 1995 Life History Correlates of Adult Size in the Malaria Vector Anopheles darlingi LP Lounibos, N Nishimura, J Conn*, R Lourenco-de-Oliveira** Florida Medical Entomology Laboratory, University of Florida, 200 9th St. SE, Vero Beach, Florida 32962, U.S.A. *Department of Biology, University of Vermont, Marsh Life Sciences Building, Burlington, Vermont 05405, U.S.A. **Departamento de Entomologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
Code Number: OC95154
Size of Files:
Text:
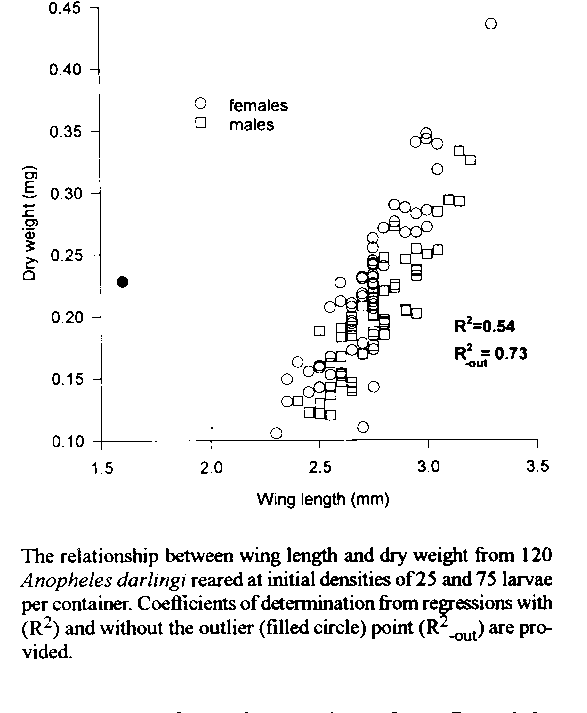
Graphics: Line Drawing (gif) - 9KAdult dry weights of laboratory-reared Anopheles darlingi were highly correlated with wing lengths, which were used to estimate size variation in natural populations of this species. Significant differences in mean wing lengths of females trapped at baits were detected among collections in the same week at one site, but not between three sites in Brazil and Bolivia. Relatively higher variability of wing lengths, compared to collections of other Anopheles (Nyssorhynchus), and platykurtic size distributions in large, single-night collections suggested that An. darlingi females caught at baits emerged from heterogenous larval habitats. No relationship was detected between parous state and the body size of wild-caught females. Adult males and females of laboratory-reared An. darlingi did not differ in body size. This absence of sexual size dimorphism is rare among mosquitoes and has not been noted previously in the genus Anopheles. Key words: Anopheles darlingi - mosquito - adult size - wing lengths - population characteristics - sexual dimorphism Since publication of the seminal paper by Fish (1985), which described natural variation in adult mosquito body sizes, many workers have noted relationships between mosquito sizes and variables of field populations such as survival (e.g., Hawley 1985, Lounibos et al. 1990), mating success (Yuval et al. 1993), fecundity (e.g., Bradshaw & Holzapfel 1992, Lyimo & Takken 1993), and infection with parasites (Lyimo & Koella 1992). Fish (1985) also suggested that properties of a mosquito populationÆs larval habitat could be deduced from the size distribution of adults trapped in nature. Only a few studies have examined size variation in natural populations of malaria vectors (Lyimo & Koella 1992, Lyimo & Takken 1993, Lounibos 1994), in spite of the awareness that vector body size distribution may affect the capacities of a population or species to transmit disease. Most, but not all (e.g., Yuval et al. 1993), body size measurements of wild-caught mosquitoes have been made on adult females. For the majority of species of mosquitoes, adult males are smaller than females and eclose earlier, a condition known as protandry (Thornhill & Alcock 1983). Protandry does not occur among some species of the mosquito genus Toxorhynchites (Corbet & Ali 1987) in which sexual size dimorphism may be absent or slight (Lounibos 1979 and unpublished data). The widespread occurrence of protandry among holometabolous insects is considered to be a reproductive strategy of males in evolutionary response to competition for mates (Wiklund & Fagerstrom 1977, Thornhill & Alcock 1983). Adult dry weight is, arguably, the most suitable indicator of size (=mass), but this measurement is tedious and often inaccessible for field studies. Many mosquito workers have, instead, measured wing lengths (e.g., Siegel et al. 1992, 1994) or pupal weights (Hawley 1985, Kleckner et al. 1995), which are both highly correlated with adult dry weights. Some early studies (e.g., Christophers 1960, Fish 1985) related mosquito dry weights to the cube of wing lengths in regression analyses, but Siegel et al. (1992) demonstrated that logarithmic data transformations provided superior fits to linear regression models. Siegel et al. (1994) cautioned against the use of wing lengths for predicting adult body weights. The principal objectives of the present study were to examine the wing length - dry weight relationship for Anopheles darlingi Root and to explore associations between body size and life history characteristics of field populations and laboratory samples of this species, the most important vector of human malaria in South America. Untransformed wing lengths were used as indicators but not as predictors of body size. Because wing lengths may underestimate the extremes of body mass (Siegel et al. 1994), our conclusions about natural size variability in An. darlingi may be conservative. MATERIALS AND METHODS Host-seeking Anopheles females were collected between 18:00 and 22:00 hr from stationary human baits at sites in Bolivia, Brazil, and Venezuela (Tables I,III). In Capanema (CP) only, An. darlingi were collected from the walls of a water buffalo corral. In Porto Velho (PV), An. darlingi were captured at human baits on three nights within one week by the same collectors in the same fazenda. Live anopheline mosquitoes held overnight in humidified containers were anesthetized and identified to species through a dissecting microscope with morphological keys (Faran & Linthicum 1981, Linthicum 1988). One wing of each specimen was removed and measured, from the base of the alula to the apical wing margin excluding fringes, with an ocular micrometer at 20X. From an arbitrarily chosen subset of An. darlingi collected at site PV, reproductive organs were dissected and parity determinations made on ovaries in Christophers' stages I or II by the tracheolar skeining technique of Detinova (1962). Interspecific comparisons of wing lengths of An. darlingi to other field-collected An. (Nysso-rhynchus) species used measurements published previously (Lounibos, 1994). The An. albitarsis (sensu lato) were identified as An. marajoara according to Linthicum (1988). Gravid An. darlingi females collected in Tefe (TE) and Puerto Ayacucho (PA) were allowed to oviposit on wet filter paper in individual vials at 26 +/-1 C in a quarantined insectary in Florida. Forty-eight hours after oviposition, eggs were floated on distilled water in plastic drinking cups. First instars hatching within 24 hr were divided into vessels containing 75 ml of distilled water and initial densities of 25 or 75 larvae. With the first day in these vessels indicated as day one, each cohort received 15 mg daily of finely ground liver powder (ICN Biochemicals), except for day three when no food was provided. Dead larvae and scum were removed daily from rearing containers. Live pupae were held individually until adult emergence in screened, nine-dram vials containing 10 ml of distilled water. Twenty-four hours after eclosion, adults were killed by freezing, sexed, and one wing was measured as described above. The effect of sex on wing lengths of An. darlingi was compared to three other species of An. (Nyssorhynchus) reared under identical laboratory conditions. Because larval density can have an effect on adult size (Lounibos 1994), mosquitoes were segregated by initial rearing density for size comparisons between the sexes. Adults reared from progeny of collections at site TE were killed by freezing, then placed in individually marked containers in a drying oven at 80 C for 48 hr. Both wings were removed for length measurements and to ensure homogeneity of weighed individuals. Oven-dried adults, maintained in a desiccator over calcium sulfate, were weighed to the nearest 0.001 mg on an electrobalance (Cahn). Analyses of variance (ANOVA) and regressions were performed on wing length and weight measurements and coefficients of variation by the GLM procedure (SAS Institute 1985). ANOVA was used, in spite of some non-normal wing length distributions, because this statistical procedure is robust to moderate departures from normality and homoscedasticity (Scheffe 1959). Following ANOVAs, means were compared by the a posteriori Ryan-Eliot-Gabriel-Welsch multiple range test (REGWQ). The relationship between wing lengths and the numbers of An. darlingi having completed one or more gonotrophic cycles was analyzed by logistic regression with the procedure CATMOD (SAS Institute 1985). Calculations and tests for significance of coefficients of variation (CV), skewness, and kurtosis followed Sokal and Braumann (1980) and Sokal and Rohlf (1981). RESULTS Relationships between dry weight and wing lengths - Regressions of dry weight (y-variate) versus wing length (x-variate) were significant both for untransformed (F1,118 = 140.25, P <0.001) and log-log transformed data (F1,118 = 95.62, P <0.001). However, the regression of untransformed data explained 9% more variance (R^2 = 0.54) than the log-transformed data (R^2 = 0.45) and, thus, only the former was subjected to further analyses (Fig.). One apparently aberrant wing length measurement of 1.60 mm was detected by the method of Sokal and Rohlf (1981) as an outlier in the regression analysis (rs = 4.60, r.005 = 3.81, n = 120, one-tailed test). When this outlier was removed, justified by the assumption that abnormal development caused the shortened wing, an added 19% variance was explained by the linear regression model (F1,117 = 317.74, P <0.001, R^2 = 0.73). Size variation in natural populations - Mean sizes of An. darlingi from CP, GU and the first night's collection at PV did not differ significantly (Table I). However, significantly different mean wing lengths were detected among the three, single-night collections of An. darlingi made within one week's span at the PV site in Brazil . Wing length CVs of mosquitoes from five night collections ranged from 5.03 to 6.70, which variability is homogeneous according to Bartlett's test performed on log-transformed variances (chi-squared = 5.41, P > 0.10). Skewnesses of wing length distributions in single-night collections were consistently negative, but none significantly so (Table I). All five wing length distributions tended to deviate platykurtically from normality, one nightÆs collection showing significantly negative kurtosis (Table I). Wing length CVs of field-collected An. darlingi were compared to CVs of four other species of An. (Nyssorhynchus) bait-trapped elsewhere in South America (Lounibos 1994). ANOVA detected significant variation among species (F4,47 = 2.77, P = 0.038), with An. darlingi recording the largest mean CV (Table II). However, a posteriori comparisons indicated no significant differences between species in mean CVs. Of 170 An. darlingi females from PV dissected for parity determinations, 104 (61.2%) were diagnosed as having completed one or more gonotrophic cycles. However, no significant relationship was detected by logistic regression between parous state and wing lengths (d.f. = 1, chi-squared = 0.63, P = 0.43, likelihood ratio = 17). Size dimorphism between sexes - The regression analyses of wing lengths versus dry weights indicated considerable weight overlap between the sexes of An. darlingi (Fig.). Male and female dry weights of An. darlingi were not significantly different among individuals reared at the low (F1,68 = 3.21, P = 0.08 ) or high density (F1,48 = 1.99, P = 0.17), nor were differences observed between wing lengths of males and females of An. darlingi from TE or PA (Table III). By contrast, sexual size dimorphism, as indicated by significant wing length differences between males and females, was pronounced among An. aquasalis, An. albitarsis (s.l.), and An. nuneztovari reared in the laboratory under the same conditions (Table III). DISCUSSION In contrast to the results of Siegel et al. (1992), who examined the fits of weight versus wing length regressions for aedine and culicine mosquitoes, untransformed data yielded a superior coefficient of determination than that obtained from the log-log transformation suggested by these authors. Our regression model for An. darlingi also explained more variance than coefficients of determination for the weight-wing length relationships of standard laboratory cultures of mosquitoes (Siegel et al. 1994). These authors showed that heterogeneous sources of mosquitoes, such as adults derived from different larval densities, may produce better regression fits than individuals reared in narrowly defined conditions. Adult An. darlingi captured in nature were smaller, on average, than three other species of the same subgenus (An. nuneztovari, An. aquasalis, and An. albitarsis), but larger than a fourth, An. triannulatus (Lounibos 1994). Among these four other species of Nyssorhynchus, no significant differences in mean wing lengths were detected among repeated night collections at the same site (Lounibos 1994), in contrast to the significant differences among captures of An. darlingi within the same week at PV. Assuming that these short-term changes in local sizes of An. darlingi reflect environmental, and not genetic, heterogeneity, the trend toward smaller mosquitoes may be related to a depletion in the quality of the larval habitat or a transition to a different primary production site of the blood-seeking adults. The mean sizes of An. darlingi females captured in the field were markedly larger than individuals reared in laboratory containers (Tables I,III), a discrepancy not observed in field versus laboratory comparisons of An. aquasalis or An. nuneztovari (Lounibos 1994). The small size of reared An. darlingi suggests that our growing conditions for this species were suboptimal. The wing length CVs of female An. darlingi from nature were larger, albeit not significantly so, than reported for the four other An. (Nyssorhynchus) examined from elsewhere in South America (Lounibos 1994). When these wing length CVs were converted to coefficients of variation for body mass (CVbm) by the method of Fish (1985), An. darlingi (CVbm = 19.5) fell into the category of moderate size variation when compared against the range of CVbms observed by Fish for North American mosquito species. The slightly higher average CVbm of An. darlingi suggests somewhat greater larval habitat variability for this species than observed for An. aquasalis (CVbm = 18.0), An. triannulatus (CVbm = 17.1), An. nuneztovari (CVbm = 15.0) or An. albitarsis (CVbm = 14.9) (Lounibos 1994). Skewness was consistently negative for all night collections of An. darlingi, although not significantly so (Table I). This pattern is consistent with other An. (Nyssorhynchus) (see Lounibos 1994) which, like An. darlingi, suffer less in nature than many temperate mosquito species from density dependent larval stresses that may produce positively skewed size distributions in natural populations (Fish, 1985) or in laboratory cultures (Bradshaw & Holzapfel 1992) of some adult mosquitoes. The uniformly negative kurtoses indicate a consistent tendency for body sizes in large collections of wild An. darlingi to cluster on either side of the mean value, and this deviation from a normal distribution was statistically significant for one nightÆs collection. The consistent platykurtic trend indicates that more females than expected are larger or smaller than the mean size, which would occur if host-seeking An. darlingi emerge from heterogeneous or diverse larval habitats. Although significant positive correlations between parous rates and body sizes have been demonstrated among diverse populations of temperate mosquitoes (e.g., Hawley 1985, Lounibos et al. 1990, Hu et al. 1993), no significant relationship was detected between wing lengths and parity in the current sample of An. darlingi, a characteristic held in common with single-night collections of An. aquasalis, An. nuneztovari, or An. triannulatus (Lounibos 1994). Although we know of no published correlations between body size and parity in a tropical anopheline, Lyimo and Takken (1993) showed that larger females of wild-caught An. gambiae (s.l.) were more likely to become gravid than smaller counterparts. The absence of sexual size dimorphism in An. darlingi is the first recognition of this phenomenon in anopheline mosquitoes. The smaller size of most male mosquitoes is typically associated with their faster development because of sex-specific selection for earlier emergence, i.e., protandry (Kleckner et al. 1995). Precise measurements of larval and pupal development times of An. darlingi remain to be done to determine whether the lack of size dimorphism in this species is associated with the absence of sex differences in time from hatch to adult eclosion.
TABLE I
Wing lengths and associated statistics of Anopheles
darlingi females captured at human baits during night
collections in Bolivia and Brazil.
Site^a Date collection Mean^b wing
Coefficient
Number length (mm) of Skewness
Kurtosis d
variation^c
GU 8-VIII-90 24 3.12A 6.15 43.34nt -0.65nt
Cp^e 20-V111-91 34 3.07AB 5.03 -0.29nt -0.40nt
PV 21-V11-94 175 3.08A 6.18 -0.17ns -0.56ns
PV 26-V1I-94 236 2.99BC 6.70 -0.09ns -0.28ns
PV 27-V11-94 220 2.96C 6.43 -0.05ns -0.71 f
a: sites - GU = Guayamerin, Beni Province, Bolivia (10 51'S,
65 21'W); CP = Capanema, State of Para, Brazil
(1 17'S,
47 34'W); PV = Porto Velho, State of Rondonia, Brazil (8^
49'S,63^ 54'W)
b: means followed by different letters are significantly
different (P <0.05) by REGWQ test
c: CV corrected for sample size
d: ns = non-significant, nt = not tested (sample size too
small)
e: mosquites trapped at animal bait
f: P< 0.05
TABLE II
Coefficients of variation^a of wing lengths of Anopheles
(Nyssorhynchus) females field-collected in South America
Species Number Number Mean S.E.
collections sites^b CV^c
--------------------------------------------------------
An. darlingi 5 3 6.10A 0.29
An. aquasalis 6 5 5.77A 0.56
An. triannulatus 10 5 5.21A 0.47
An. albitarsis (s.l) 11 5 4.82A
0.33
An. nuneztovari 20 9 4.73A 0.12
a: CVs corrected for sample size b: specific
site information available in Lounibos (1994) c: means
followed by different letters are significantly (P <0.05)
different by REGWQ test
TABLE III
Mean wig length of Anopheles darlingi separated by sex and of
three species of the same subgenus reared in standard
laboratory conditions
--------------------------------------------------------------
Males Females
Anopheles sp. ------------------- ------------------------
Mean Mean
Origin Init. No. w.l. SE No. w.l. SE p^b
of larval (mm) (mm)
(site) dens
^a
--------------------------------------------------------------
darlingi
TE 25 34 2.81 0.03 36 2.77 0.03 0.377
darlingi
PA 25 90 2.63 0.01 73 2.62 0.01 0.581
aquasalis
SF 25 71 3.11 0.02 51 3.33 0.02 <0.001
albitaris (s.l.)
RS 25 49 3.00 0.02 48 3.14 0.02 <0.001
mineztovari
RS 75 60 2.86 0.02 72 2.96 0.02 0.007
^a: TE = Tefe State of Amazonas, Brazil (3o22'S, 64o43'W); PA
= Puerto Ayacucho, State of Amazonas, Venezuela (5o59'N,
67o39'W); SF = Santa Fe, STate of Sucre, Venezuela
(10o07'N,64o25'W); RS = Rio Socuavo, STate of Zulia, Venezuela
(8o54'N,72o38'W)
^b: probabilities from unpaired t-test (two-tailed) comparing
male and female wing length.
ACKNOWLEDGEMENTS
To staff of Fundacao Nacional da Saúde (Brazil), especially F dos Santos, CAB Queiroz, J Artiago, HP Nunes Filho, K Marins de Moraes and A Cruz Marques; Servicio Nacional de la Eraddicacion de Malaria (Bolivia), especially J Cuba, C Toffen, and A Melgar; Direccion de Malariologia y Saneamineto Ambiental (Venezuela), especially J Mora and J Toro, for their help in field collections of An. darlingi; to D Couto Lima of FIOCRUZ for performing dissections; to R Zimmerman, B Yuval, J Rey and G O'Meara for comments on an earlier draft.
REFERENCES
Bradshaw WE, Holzapfel CM 1992. Reproductive consequences of density-dependent size variation in the pitcher plant mosquito, Wyeomyia smithii (Diptera: Culicidae). Ann Ent Soc Amer 85: 274-281. Corbet PS, Ali HJ 1987. Diel patterns of pupation and emergence, and protogyny, in Toxorhynchites brevipalpis brevipalpis (Theobald) (Diptera: Culicidae): a laboratory study. Ann Trop Med Parasit 81: 437-443. Christophers SR 1960. Aedes aegypti (L.): the Yellow Fever Mosquito - its life history, bionomics, and structure. Cambridge University Press, xii + 739pp. Detinova RS 1962. Age grouping methods in Diptera of medical importance. WHO, Monograph 47. Faran ME, Linthicum KJ 1981. A handbook of the Amazonian species of Anopheles (Nyssorhynchus) (Diptera: Culicidae). Mosq Syst 13:1-81. Fish D 1985. An analysis of adult size variation within natural mosquito populations, p. 419-429. In LP Lounibos, JR Rey, JH Frank (eds), Ecology of Mosquitoes: Proceedings of a Workshop. Florida Medical Entomology Laboratory, Vero Beach, Florida, xxii + 579pp. Hawley WA 1985. Population dynamics of Aedes sierrensis, p. 167-184. In LP Lounibos, JR Rey, JH Frank (eds), Ecology of Mosquitoes: Proceedings of a Workshop. Florida Medical Entomology Laboratory, Vero Beach, Florida, xxii + 579pp. Hu GY, Lounibos LP, Escher RL 1993. Seasonal abundance, age composition and body size of salt marsh Anopheles (Diptera: Culicidae) in south Florida. J Med Ent 30: 883-887. Kleckner CA, Hawley WA, Bradshaw WE, Holzapfel CM, Fisher IJ 1995. Protandry in Aedes sierrensis: the significance of temporal variation in female fecundity. Ecology 76: 1242-1250. Linthicum KJ 1988. A revison of the Argyritarsis Section of the subgenus Nyssorhynchus of Anopheles. Mosq Syst 20: 98-271. Lounibos LP 1979. Temporal and spatial distribution, growth and predatory behaviour of Toxorhynchites brevipalpis on the Kenya coast. J Anim Ecol 48: 213-236. Lounibos LP 1994. Geographical and developmental components of adult size of neotropical Anopheles (Nyssorhynchus). Ecol Ent 19: 138-146. Lounibos LP, Larson VL, Morris CD 1990. Parity, fecundity and body size of Mansonia dyari in Florida. J Am Mosq Cont Assoc 6: 121-126. Lyimo EO, Koella JC 1992. Relationship between body size of adult Anopheles gambiae s.l. and infection with the malaria parasite Plasmodium falciparum. Parasitol 104:233-237. Lyimo EO, Takken W 1993. Effects of adult body size on fecundity and the pre-gravid rate of Anopheles gambiae females in Tanzania. Med Vet Entomol 7: 328-332. SAS Institute Inc. 1985. Sas UserÆs Guide. Statistics. Version 5 ed. Cary, North Carolina, xvi+956pp. Scheffe H 1959. The Analysis of Variance. John Wiley & Sons, New York. Siegel JP, Novak RJ, Lampman RL, Steinley BA 1992. Statistical appraisal of the weight-wing length relationship of mosquitoes. J Med Ent 29: 711-714. Siegel JP, Novak RJ, Ruesink WG 1994. Relationship between wing length and dry weight of mosquitoes. J Am Mosq Control Assoc 10: 186-192. Sokal RR, Braumann CA 1980. Significance tests for coefficients of variation and variability profiles. Syst Zool 29: 50-66. Sokal RR, Rohlf FJ 1981. Biometry. 2nd ed. W H Freeman, San Francisco, xviii + 859 pp. Thornhill R, Alcock J 1983. The Evolution of Insect Mating Systems. Harvard University Press, Cambridge, Massachusetts, ix + 547pp. Wiklund C, Fagerstrom T 1977. Why do males emerge before females? A hypothesis to explain the incidence of protandry in butterflies. Oecologia 31: 153-158. Yuval B, Wekesa JW, Washino RK 1993. Effect of body size on swarming behavior and mating success of male Anopheles freeborni (Diptera: Culicidae). J Ins Behav 6: 333-342. This work was supported by grants from the National Institutes of Health (AI-31034) of the U.S.A. and CNPq, PAPES/FIOCRUZ and FNS of Brazil, and is University of Florida IFAS Journal Series No. R-04434.
Received 7 March 1995 Accepted 1 August 1995 Copyright 1995 Fundacao Oswaldo Cruz, FIOCRUZ
The following images related to this document are available:Line drawing images[oc95154a.gif] |
| |||||||||

{kind=link}