 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
N-Terminal Amino Acid Sequences of the Major Outer Membrane Proteins from a Neisseria meningitidis Group B Strain Isolated in Brazil Salvatore Giovanni De Simone/+/**, Solange AF Soares*, Andre LA Souza*, Maria GM Danelli* Laboratorio de Microsequenciamento de Proteinas, Departamento de Bioquimica e Biologia Molecular, Instituto Oswaldo Cruz *Laboratorio de Vacinas Bacterianas, Departamento de Desenvolvimento Tecnologico, Instituto de Tecnologia - FIOCRUZ, Av. Brasil 4365, 21040-900 Rio de Janeiro, RJ, Brasil **Departamento de Biologia Celular e Molecular, Instituto de Biologia, Niteroi, Rio de Janeiro, Brasil
Code Number: OC96020
Sizes of Files:
Text: 27K
Graphics: Photos (jpg) 55K
This research was supported by CNPq (No 50.0445/92.3) and FIOCRUZ +Corresponding author: FAX: 055 (021) 590-3495; E-mail: dsimone@gene.dbbm.fiocruz.br Received 7 March 1995 Accepted 8 November 1995 The four dominant outer membrane proteins (46, 38, 33 and 28 kDa) were detected by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) in a semi-purified preparation of vesicle membranes of a Neisseria meningitidis (N44/89, B:4:P1.15:P5.5,7) strain isolated in Brazil. The N-terminal amino acid sequence for the 46 kDa and 28 kDa proteins matched that reported by others for class 1 and 5 proteins respectively, whereas the sequence (25 amino acids) for the 38 kDa (class 3) protein was similar to class 1 meningococcal proteins. The sequence for the 33 kDa (class 4) was unique and not homologous to any known protein.
Key words: Neisseria meningitidis group B - outer membrane proteins - class specific proteins - N-terminal amino acid sequence Within the past decade, groups of investigators have begun to characterize on a molecular level the structural components of Neisseria meningitidis, the ethiologic agent of meningococcal disease. Many of these investigators (Tsai et al. 1981, Achtman et al. 1983, Morse et al. 1983, Poolman et al. 1983, Frasch et al. 1986, Barnejee-Bhatnagar & Frasch 1990) have sought to identify and to analyze several meningococcal surface components in an attempt to define important immunogens or potential vaccinant compounds. Unfortunatedly, for the human disease caused by the meningococci serogroup B, the capsular polysaccharide (CP) has shown poor immunogenicity (Wyle et al. 1972) and in some cases induced immunological tolerance (Frasch 1990, Poolman 1995). A likely explanation for this last property of CP is its structural identity, a homopolymer of a2®8 sialic acid, with polysialyated host glycoproteins (Finne et al. 1987). The N. meningitidis serogroup B is the predominant etiologic agent of the infection in many countries (Whiattle & Greenwood 1976, Bower et al. 1977, Peltora 1985, Cougant et al. 1986), and specifically in Brazil it has been reported that the number of cases is increasing since 1976 (Sacchi et al. 1992).
Current experimental vaccines are therefore based on the outer membranes compounds (Poolman 1995) which have been treated to remove the potentially toxic lipopolysaccharide (LPS) (Poolman et al. 1983, Cowe et al. 1989, Banerjee-Bhatnagar & Frash 1990). Such preparations contain variable amounts of five classes of major outer membrane proteins (OMP), namely C1 through C5, which have apparent molecular weights of 46.000, 41.000, 38.000, 33.000 and 28.000, respectively (Tsai et al. 1981, Van der Ley et al. 1991, 1993 a,b). C1 (class 1 OMP) is a porin-like molecule and is generally present in most meningococcal strains. The antigenic variations of this protein define the subtype of a meningococcal strain (Poolman 1995) and to date, monoclonal antibodies (MAbs) have been made against 15 different subtypes (Abdillahi & Poolman 1988). As C1, the C2 (class 2 OMP) and C3 (class 3 OMP) are porin-like proteins (anion-specific) and are mutually exclusive since meningococcal strains have either the class 2 or class 3 proteins. These antigens represent the predominant proteins on the outer membrane (Tommassen et al. 1990, Van der Ley et al. 1991, Van der Ley & Poolman 1992) and, because of their limited antigenic variation, they have been used to define at least 20 serotypes (Griffiss et al. 1977). C4 (class 4 OMP) appears to be the highly conserved between meningococcal strains. Although its cellular function is unknown, it shares sequence homology with Escherichia coli Omp A (Klugman et al. 1989). Antibodies directed against this protein are non-bactericidal and have the additional ability to block the lytic effect of antibodies directed against other outer surface antigens (Munkley et al. 1991). Finally, C5 (class 5 OMP, opc or opacity-associated protein), in contrast to the other major OMP, is quite variable both in its qualitative and quantitative expression (Achtman et al. 1991, Aho et al. 1991, Poolman 1995) but may induce bactericidal antibodies (Danelli et al. 1995). This group of proteins also elicits strong but strain-specific antibody responses and confers important interstrain antigenic differences which may be detected by MAbs (Zollinger et al. 1984). As some of these proteins are targets for bactericidal antibodies and show characteristic differences it was decided to determine the primary sequence of the class 1-5 proteins. Advances in the structural studies of OMPs not only contributed significantly to the understanding the structure-function relationships, but made it possible to characterize conformational changes which accompany the reajustments of amino acid residues involved in the immune response of the host. In this work we describe the N-terminal amino acid sequence of the class 1-5 N. meningitidis OMPs from a strain recently isolated in Sao Paulo, Brazil. The data can contribute to the identification of important antigenic determinants and the elucidation of the structural and functional properties of the molecules.
MATERIALS AND METHODS Bacterial strain and obtention of the outer membrane proteins- N. meningitidis N44/89 (B:4:P1.15:P5.5,7) was isolated from a patient with meningococcal septicemia by the Bacteriological Division of the Adolfo Lutz Institute, Sao Paulo, Brazil. The outer membrane proteins were prepared as described previously (Frasch 1990) with some modifications. Briefly, the bacterium was cultured in Catlin's medium and the outer membrane vesicles (OMVs) were obtained by centrifugation (3.000g, 1hr, 25 C). The pellet was ressuspended in 0.1M Tris-HCl buffer, pH 8.5, containing 0.2 mM EDTA (10% w/v, Tris-EDTA buffer) and the suspension sheared in an ice cooled, omnimixer (Sorvall) for 10 min. The cells were removed by two successive centrifugation steps at 10.000g and 12.000g for 20 min each. The OMVs in the supernatant were then pelleted at 37.000g for 45 min, resuspended in Tris-EDTA buffer and stored at -20 C until use. SDS-PAGE and protein determination - The OMVs proteins were analyzed by polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate (SDS-PAGE, Laemmli 1970) under reducing conditions and the protein concentration was measured according to Lowry's method (Lowry et al. 1951), using bovine serum albumine as a standard. N-terminal amino acid sequence analysis - For sequencing experiments, the isolated proteins were run on a 15%-acrylamide SDS-PAGE (Laemmli 1970) and the proteins blotted on to a polyvinylidene difluoride (PVDF) membrane (Bio-Rad Laboratories Ltd, UK) using a semi-dry blotting apparatus (Bio-Rad). The partial amino N-terminal sequences were performed by automatic Edman degradation using a gas-phase protein microsequencer (model PSQ-1, Shimadzu) coupled to an on-line high-performance liquid chromatography (model 6A; Shimadzu, Kyoto, Japan), as described previously (Giovanni De Simone et al. 1994). The initial yield was 55-65% and the repetitive yield 90-92%. A Digital Decpc computer was used to store the GenBank, EMBL and PIR databases. The nucleotide and deduced amino acid sequences were used to query these databases using Wiscousin GCG FASTA algorithmus.
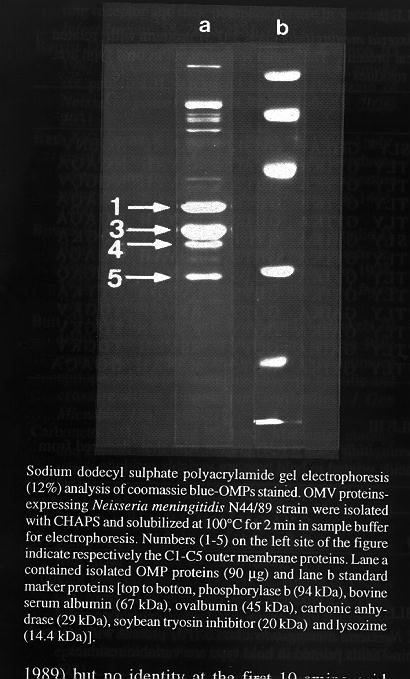
RESULTS AND DISCUSSION The SDS-PAGE analysis of the isolated OMPs (N. meningitidis serogroup B) is shown in Fig. The Mr of the identified proteins are in good accordance with those from the meningococcal described in the literature. Amino acid analysis of C1, C3, C4 and C5 proteins showed that none of them presented identical N-terminal sequences with the possible exception of the C1 and C3. These two proteins were homologous (40% identity in 25 amino acids) but clearly different in the positions 3,7,13-25. The common sequence D1 V2 S3 L4 Y5 G6 I8 K9 A10 G11 V12 was identical for the first 12 amino acids to the sequence predicted from DNA sequencing of different genes (position 20-39) for meningococcal serogroup B and C class 1 (Table I) and class 3 precursor proteins (Table II). A high similarity with PIA and PIB gonococcal proteins also was observed.
The N-terminal amino acid sequence of the class 4 protein begins with the glycine residue and is distinct from the class 1 and 3 proteins but similar (50% in 10 amino acids) to a class 4 serotype 2a deduced meningococcal sequence (Table III). The amino acid sequence similarity of gonococcal protein III with the OmpA protein of E. coli and other enterobacteria has been described elsewhere (Gotschlich et al. 1987, Klugman et al. 1989) but no identity at the first 10 amino acid appears with the meningococcal class 4 protein determined in this study. The N-terminal amino acid sequence of the C5 protein presented a high similarity (60%) with a class 5 serogroup A meningococcal protein (Achtman et al. 1983) isolated from patients during the Gambia (West Africa) epidemic meningococcal meningitis (clone A IV-1) in 1982-1983. A similarity of 100% with a C5a (heat modified form) isolated protein was observed with a translated DNA sequence (pFLOB540) from FAM18 cells (Kawula et al. 1988) but no identity was see with the PII gonococcal protein (Table IV). TABLE I Comparison of N-terminal amino acid sequence of Neisseria meningitidis class 1 (C1) protein with deduced and translated meningococcal class 1 proteins. The amino acids printed in bold type are variable residues
------------------------------------------------------------
Strain Serotype Gen bank accession N-terminal sequence
no./Refence
------------------------------------------------------------
N44/89 B:4P1.15 This work DVSLY^5 GEIKA^10 GVQGR^15
NFQLE^20 IEPP^25
H44/76 B:15P1.7.1 McGuinness DVSLY GEIKA GVEGR
et al. 1990 NYQLQ LTEAQ
MC50 C:NT:P1.16 Barlow et DVSLY GEIKA GVEGR
al. 1990 NIQAQ LTEQP
MC51 C:NT:P1.15 McGuinness DVSLY GEIKA GVEGR
et al. 1990 NFQLQ LTEPP
2996 B:2b:1.2 X60105 DVSLY GEIKA GVEGR
NIQLQ LTEPL
------------------------------------------------------------
NT: not typedTABLE II Alignment of N-terminal amino acid sequence of Neisseria meningitidis class 3 (P3) proteins with related class 2 and 3 translated meningococcal and gonococcal proteins. The amino acids printed in bold type are variable residues
------------------------------------------------------------
Strain Serotype Gen bank accession N-terminal sequence
no./Refence
------------------------------------------------------------
N44/89 B:4P1.15 This work DVSLY^5 GEIKA^10 GVEIKI^15
RKVKI^20 NGENV^25
12GE 3 U07191 DVTLY GTIKA GEATS
RSVAH NGAQA
BB1350 4 U07193 DVTLY GTIKA GETS
RSVEH NGGQV
M978 8 U07189 DVTLY GTIKA GETS
RSVAH NGAQA
S3446 14 U07188 DVTLY GTIKA GETS
RSVAH NGAQA
6557 17 U07190 DVSLY GEIKA GVETS
RSVYY EDGKA
1901 18 U07192 DVTLY GTIKA GETS
RSVAH NGAQA
6940 19 U11030 DVTLY GTIKA GETS
RSVAH NGGAQ
M986 B:2a Murakami et DVTLY GTIKA GEVS
al. 1989 RVVDA GTYKA
P9 PIB Carbonetti et DVTLY GAIKA GQTY
al. 1987 RSVEH TDGKV
MSII PIB Gotschlich et DVTLY GAIKA GQTY
al. 1987 RSVEH TDGKV
RIO PIB Butt et DVTLY GAIKA GQTY
al. 1990 RSVEH TDGKV
FA19 PIA Carbonetti et DVTLY GTIKA GETS
al. 1988 RSVAH HGAQA
-------------------------------------------------------------
TABLE IIIAlignment of aminoterminal sequence of Neisseria meningitidis class 4 (C4) protein with that deduced from DNA meningococcal sequence. The amino acids printed in bold type are variable residues
-------------------------------------------------------- Strain Serotype Reference N-terminal sequence -------------------------------------------------------- N44/89 B:4P1.15 This work GVAQV^5NHYKV^10 M986 B:2a Klugman et al. 1989 GEASVQGYTV --------------------------------------------------------TABLE IV Comparison of the N-terminal amino acid sequence of Neisseria meningitidis class 5 (P5) protein with related meningococcal and gonococcal proteins. The amino acids printed in bold type are variable residues
---------------------------------------------------------- Strain Serotype Reference N-terminal sequence ---------------------------------------------------------- N44/89 B:4P1.15 This work ASELG^5 s RSPY^10 pFLOB540 Kawula et at. 1988 AsELG RSPY FAM18 C2a Achtman et al. 1983 AsEGs sGrPY FA1090 PH Bamtt et al. 1987 AEGRP YVNAD ---------------------------------------------------------- Although a very high homology between the class 1 (20 amino acids) and class 5 proteins (10 amino acids) of the N. meningitidis serogroup B was observed with similar proteins from serogroup A described in the literature, previous immunological characterization of the strains showed that they present different antigenic properties. Moreover, the sequence of amino acids 1 through 20, of the class 1 and 3 proteins, has been suggested to comprise a highly conserved transmembrane protein segment of Neisseria sp. while the variable segment seem to be at the outside of the membrane forming an important antigenic site (McGuiness et al. 1990, Bash et al. 1995). Thus it is important go on in the primary structure of these proteins in order to define their variable regions. The class 1 protein is unique to meningococci, it is immunogenic and antibodies directed against it are bactericidal in vitro (Frasch et al. 1986, Poolman et al. 1987) and protect against infection in an experimental animal model (Saukkonen et al. 1987). The antigenic differences in the class 2/3 outer membrane proteins are important and accountable for serotype specificity, while antigenic determinants on the class 1 protein generate subtype specificity. A recent study, using synthetic peptides from three strains of N. meningitidis (MC50, MC51, H44/76) defined the regions VR1 (24-36) and VR2 (179-187) from the class 1 proteins as responsible for the serosubtype specificity (McGuinness et al. 1990). The determination of the structure-function relationship can provide important clues for the understanding of the multiple biological effects of these outer membrane proteins on different strains of N. meningitidis. In addition, the DNA sequence or selected constituent oligonucleotide portions, including synthetic oligonucleotides, may be used as molecular gene probes for the detection of the organism in various body fluids. Knowledge of the amino acid sequence also allows the testing of strategic synthetic peptides to identify and confirm immunodominant B-cell or T-cell epitopes. Synthetic peptides also may be used as the basis for improve N. meningitidis serologic tests and/or N. meningitidis synthetic peptide vaccines.
REFERENCES Abdillahi H, Poolman JT 1988. Neisseria meningitidis group B serosubtype using monoclonal antibodies in whole cell ELISA. Microb Pathog 4: 27-32. Achtman M, Neibert M, Crowe BA, Strittmatter W, Kusecek B, Weyse E, Walsh M, Slawing B, Morelli G, Moll A, Blake M 1983. Purification and characterization of eight class 5 outer membrane protein variants from a clone of Neisseria meningitidis serogroup. Am J Exp Med 168: 507-525. Achtman M, Wall RA, Bopp M 1991. Variation in class 5 protein expression by serogroup A meningococci during a meningitidis epidemic. J Infect Dis 164:375-382. Aho EL, Dempsey JA, Hobbs MM, Klapper DG, Cannon JG 1991. Characterization of the opA (class 5) gene family of Neisseria meningitidis. Mol Microbiol 5: 1429-1437. Barlow AK, Heckels JE, Clarke IN 1989. The class 1 outer membrane protein of Neisseria meningitidis: gene sequence and structural and immunological similarities to gonococcal porins. Mol Microbiol 3: 131-139. Barnejee-Bhatnagar N, Frasch CE 1990. Expression of Neisseria meningitidis Iron-regulated outer membranes proteins, including a 70 Kilodalton transferring receptor and their potential for use in vaccines. Infect Immun 58: 2875-2881. Barritt DS, Schwalbe RS, Klapper DG, Cannon JG 1987. Antigenic and structural differences among six proteins II expressed by a single strain of Neisseria gonorrhoeae. Infect Immun 55: 2026-2031. Bash MC, Lesiak KB, Banks S, Frash CE 1995. Analysis of Neisseria meningitidis class 3 outer membrane protein gene variable region and type identification using genetic thechniques. Infect Immun 63: 1484-1490. Bower K, Holten E, Vik-mo H, Brondbo A, Bratlid D, Bjark P, Moe PJ 1977. Neisseria meningitidis infections in Northern Norway: An epidemic in 1974-1975 due mainly to group B organisms. J Infect Dis 135: 669-672. Butt NJ, Virji M, Vayreda F, Lambden PR, Heckels JE 1990. Gonococcal outer membrane protein PIB: comparative sequence analysis and localization of epitopes which are recognized by type-specific and cross-reacting monoclonal antibodies. J Gen Microbiol 136: 2165-2172. Carbonetti NH, Sparling PF 1987. Molecular cloning and characterization of the structural gene for protein I, the major puter membrane protein of Neisseria gonorrhoeae. Proc Natl Acad Si USA 84: 9084-9098. Carbonetti NH, Smnad VI, Seifer HS, So M, Sparling PF 1988. Genetics of protein I of Neisseria gonorrhoeae: construction of hybrid porins. Proc Natl Acad Sci USA 85: 6841-6845. Cougant DA, Frohon LO, Bouvre K, Holten E, Frasch CE, Mocca LF, Zollinger WD, Selander LK 1986. Intercontinental apread of a genetically distinctive complex of clone of Neisseria meningitidis causing epidemic disease. Proc Natl Acad Sci USA 83:4927-4931. Cowe BA, Wall RA, Kuseck B, Newman B, Olyhock T, Achtman M 1989. Clonal and variable properties of Neisseria meningitidis isolated from cases and carries durin and after an epidemic in the Gambia, West Africa. J Infect Dis 159: 686-700. Danelli MG, Batoreu NM , Lacerda MD, Ferreira CRB, Cardoso JD, Peralta JM, Frasch CE 1995. Surface antigen analysis of group B Neisseria meningitidis outer membranes by monoclonal antibodies identification antibodies Ac 5 protein. Current Microbiol 31: 146-151. Finne J, Bitter-Sverman D, Goridis C, Finne U 1987. An IgG monoclonal antibody to group B meningococci cross-reacts with developmentally regulated polysialic acid units of glycoproteins in neural and extraneural tissues. J Immunol 138: 4402-4407. Frasch CE 1990. Production and control of Neisseria meningitidis vaccines, p. 123-145. In Mizrahi A, Bacterial Vaccines. John Willey Sens Inc, New York. Frasch CE, Tsai C, Mocca LF 1986. Outer membrane proteins of Neisseria meningitidis: Structure and importance in memingococcal disease. Clin Invest Med 9: 101-108. Giovanni De Simone S, Santos R, Araujo MF, Pinho RT 1994. Preparative isolation of the lectin jacalin by anion exchange high-performance liquid chromatography. J Chromatography A 688: 357-362. Gotschlich EC, Seiff M, Blake MS 1987. The DNA sequence of the structural gene of gonococcal protein III and the flanking region containing a repetitive sequence. Homology of protein III with enterobacterial OmpA proteins. J Exp Med 165: 471-482. Griffiss ML, Broud DD, Silver CA, Artenstein MS 1977. Immunoepidemiology of meningococcal disease in military recruits. A model for serogroup independency of epidemic potential as determined by serotyping. J Infect Dis 136: 176-186. Kawula TH, Aho EL, Barritt DS, Klapper DG, Cannon JG 1988. Reversible phase variation of expression of Neisseria meningitidis class 5 outer membrane proteins and their relationship to gonococcal proteins II. Infect Immun 56: 380-386. Klugman KP, Gotschlich EC, Blacke MS 1989. Sequence of the structural gene ( rmp M ) for the class 4 outer membrane protein of Neisseria meningitidis homology of the protein to gonococcal protein III and Escherichia coli OMP 4 and construction of meningococcal strains that lock class 4 protein. Infect Immunol 57: 2066-2071. Laemmli UK 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685. Lowry OH, Rosebrugh NJ, Farr AL, Randall RJ 1951. Protein measurement with the folin phenol reagent. J Biol Chem 193: 265-275. McGuinness B, Barlow AK, Clarke IA, Farley JE, Anilionis A, Poolman JT, Heckels JE 1990. Deduced amino acid sequences of class 1 protein (Por A) from three strains of Neisseria meningitidis. J Exp Med 171: 1871-1882. Morse SA, Mintz CS, Sarafian SK, Bartenstein M, Bertram M, Apicella MA 1983. Effect of dilution rate on lipopolysaccharide and serum resistance of Neisseria gonorrhoeae grown in continuous culture. Infect Immun 41: 74-82. Munkley A, Tinsley CR, Virji M, Heckels JE 1991. Blooting of bactericidal killing of Neisseria meningitidis by antibodies directed against class 4 outer memmbranes proteins. Microbiol Pathol 11: 447-452. Murakami K, Gotschlich EC, Seiff ME 1989. Cloning and characterization of the structural gene for the class 2 protein of Neisseria meningitidis. Infect Immun 57: 2318-2323. Peltora J 1985. Meningococcal disease: still with us. Rev Infect Dis 7: 504-511. Poolman JT 1995. Development of a meningococcal vaccine. Infect Immun 4: 13-28. Poolman JT, Hopman CTP, Zanan HC 1983. Immunogenicity of meningococcal antigens as detected in patients sera. Infect Immun 41: 398-406. Poolman JT, Timmermans AM, Hopman CTP, Teerlink T, Van Vught PAM, Witvliet MH, Beuvery EC 1987. Comparison of meningococcal outer membrane proteins vaccines solubilized with detergent or C polysaccharide. Antonie Leeuwenhoek J Microbiol 53: 261-266. Sacchi CT, Pessoa LL, Ramos SR, Milagres LG, Camargo MCC, Hidalgo NTR, Melles CEA, Caugant DA, Frasch CE 1992. Ongoing group B Neisseria meningitidis epidemic in Sao Paulo, Brazil, due to increased prevalence of a single clone of the ET.5 complex. J Clin Microbiol 30: 1734-1738. Saukkonen K, Abdillahi H, Poolman JT, Lenonen M 1987. Protective efficacy of monoclonal antibodies to class 1 and class 3 outer membrane proteins of Neisseria meningitidis B:15:P1.16 in infant rat infection model. New perpects for vacine development. Microb Pathog 3: 261-269. Tommassen J, Verney P, Struyve M, Bens R, Poolman JT 1990. Isolation of mutants of Neisseria meningitidis deficient in class 1 (por A) and class 3 (por B). Infect Immun 58: 1355-1359. Tsai CM, Frasch CE, Mocca LF 1981. Five structural class of outer membrane proteins of Neisseria and Neisseria meningitidis. J Bacteriol 146: 69-74. Van der Ley PA , Heckels JE, Virji M, Hoogerhout P, Poolman JT 1991. Topology of outer membranes porins in pathogenic Neisseria spp. Infect Immun 59:296-271. Van der Ley PA, Van der Biezen J, Peeters CCAM, Poolman JT 1993a. Use of transformation to construct antigenic hibrids of the class 1 outer membranes protein in Neisseria meningitidis. Infect Immun 61: 4724-4733. Van der Ley PA, Van der Biezen J, Poolman JT 1993b. Construction of Neisseria meningitidis strains multiple cromossomal copies of the por A gene for use in the production of a multivalent outer membrane protein vaccine. Vaccine (in press). Whiattle HC, Greenwood BM 1976. Meningococcal meningitis in the Northem savana of Africa. Trop Doct 6: 99-104 . Wyle FA, Artenstein MS, Brandt BL, Tramont EC, Kasper DL, Altieri PL, Bernam SL , Lowented JL 1972. Immunological response of may to group B meningococcal polissacharides vaccines. J Infect Dis 126: 514 -522. Zollinger W, Moran EE, Connelly H, Mandrell R, Brandt B 1984. Monoclonal antibodies to serotype 2 and serotype 15 outer membrane proteins of Neisseria meningitidis and their use in serotyping. Infect Immun 46: 260-266.
Copyright 1995 Fundacao Oswaldo Cruz
The following images related to this document are available:Photo images[oc96020a.jpg] |
| |||||||||

{kind=link}