 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Trypanosoma cruzi Infection in Offspring Born to Chagasic C^3H/He Mice
Silvana Marques de Araujo^+, Egler Chiari*
Setor de Parasitologia Basica, Universidade Estadual de Maringa, Caixa Postal 331, 87020-900 Maringa, PR, Brasil *Departamento de Parasitologia, ICB/UFMG, Caixa Postal 2486, 31270-901 Belo Horizonte, MG, Brasil
Code Number: OC96042
Size of Files:
Text: 24K
Graphics: Line Drawing (gif) - 18.4K[TABLE AND FIGURE AT END OF TEXT] This study reports the effects of Trypanosoma cruzi infection induced in C3H/He male and female mice born to chagasic mice. An experimental model was established infecting female C3H/He mice with a low virulent T. cruzi clone. In this model, mating, fertilization, pregnancy evolution and delivery was carried out successfully. The offspring was infected at four, six and eigth weeks of age. The results showed that the offspring born to chagasic mothers present decreased resistance to acquired T. cruzi infection. This decreased resistance was expressed by higher levels of parasitaemia and higher mortality rates in offspring born to chagasic mothers than in controls. Age and sex were shown to be important factors of this phenomenon. The results suggest that maternal immune system products can modulate the immune response of the offspring.
Human chagasic infection is widespread in tropical and neotropical regions where there are triatominae bugs, vertebrate reservoirs and where ecological conditions favor the dissemination of Trypanosoma cruzi (Hoare 1972). Serological tests have shown that the prevalence of chagasic human infection ranges from 0.43% in Georgia, USA (PAHO 1970) to 74.2% in hyperendemic areas like some in Argentina (Cichero et al. 1967). Among pregnant women, it ranges from 2 to 51% in urban areas (Passos 1960, Espinal 1962, Azougue et al. 1985) and from 23 to 81% in endemic areas (Barousse et al. 1978, Lugones et al. 1979, Azougue et al. 1985). Due to these endemic characteristics many individuals who acquired or are at risk of acquiring Chagas' disease certainly were exposed to parasite antigens and/or to anti-T. cruzi antibodies from their mothers during pregnancy and breast-feeding. Experimental Chagas' disease in mice induce a polyclonal activation of B and T lymphocytes (Corsini & Costa 1981, D'Imperio Lima et al. 1985, Minoprio et al. 1986 a,b). The transfer of maternal antibodies, antigens, immune complex and cytokines through placental or breast-feeding significantly affects the offsprings' immune system. This phenomenon possibly results in a change of idiotypic connectivity patterns in the developing immune system (Wikler et al. 1980, Nisonoff et al. 1983). The present study reports the effects of T. cruzi infection induced at different ages in male and female mice born to chagasic female mice using parameters involved in biological evaluation of experimental chagasic infection.
MATERIALS AND METHODS
Animals and mating - Male and female C3H/He mice were used in the experiments. Eight week old animals were used for mating and four, six and eight week old animals were used for experimental evaluation. These animals were provided by the Department of Parasitology, University of Minas Gerais, Belo Horizonte, MG, Brasil. Females were mated 75 days after T. cruzi infection. For mating two to four females were housed in a cage containing only one male. Animals were kept separated until mating. Control groups were subject to the same procedure. T. cruzi infection - The female mice for the mating were injected intraperitoneally with 500 bloodstream trypomastigotes from a clone of Y strain, called YP1 (Araujo & Chiari 1988). In order to determine the parasitaemia patterns and the mortality rates, groups of animals born to infected female mice mothers were inoculated with 5,000 trypomastigotes. Birth evaluation - Birth date, litter size, weight and survival rate were determined by a daily visit to the animal room until weaning at four weeks. Comparisons between infected and control groups were made by the Student's t test. Congenital transmission - Four week old surviving animals born to chagasic mothers were bled by orbital puncture on the weaning day and the blood was cultured in two different tubes containg 3ml of liver infusion trypyose (LIT) medium (Camargo 1964). Aliquots of 10 ul from each tube were examined every 15 days up to 45 days after inoculation. Mice born to control mothers were bled in order to cause the same stress. In previous experiments the efficiency of this method for parasite detection was 93.3%. T. cruzi infection evaluation - (a) Parasitaemia - Groups of five to eight male or female mice, four, six and eight week old, born to chagasic or control mothers were used to determine the parasitaemia. Parasitaemia curves were obtained by daily examination from 4 to 30 days after infection. The number of parasites was established by the Brener's technique (Brener 1962). Data shown are arithmetic mean of the animals used in each group. Comparisons between parasitaemia obtained from mice born to chagasic mothers and born to control groups were made by the Student's t test. (b) Mortality - Groups of 6 to 13 animals born to chagasic or normal mothers were observed for 120 days after inoculation and their deaths registered daily. The results were expressed in percentage of cumulative mortality per sex/age during the observation period. Results were compared by the Chi square test.
RESULTS
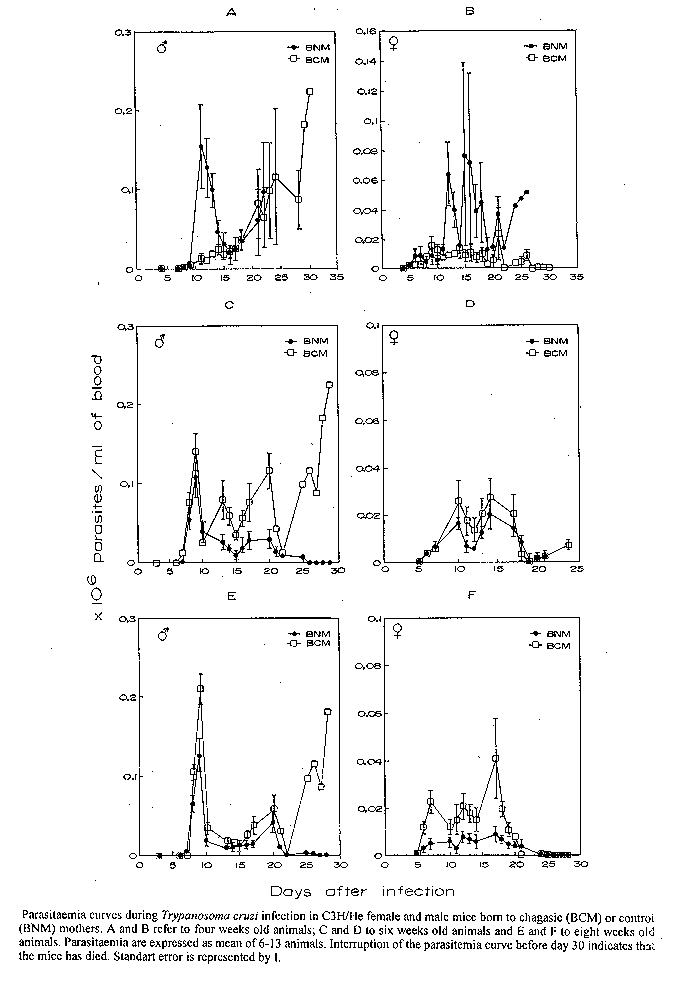
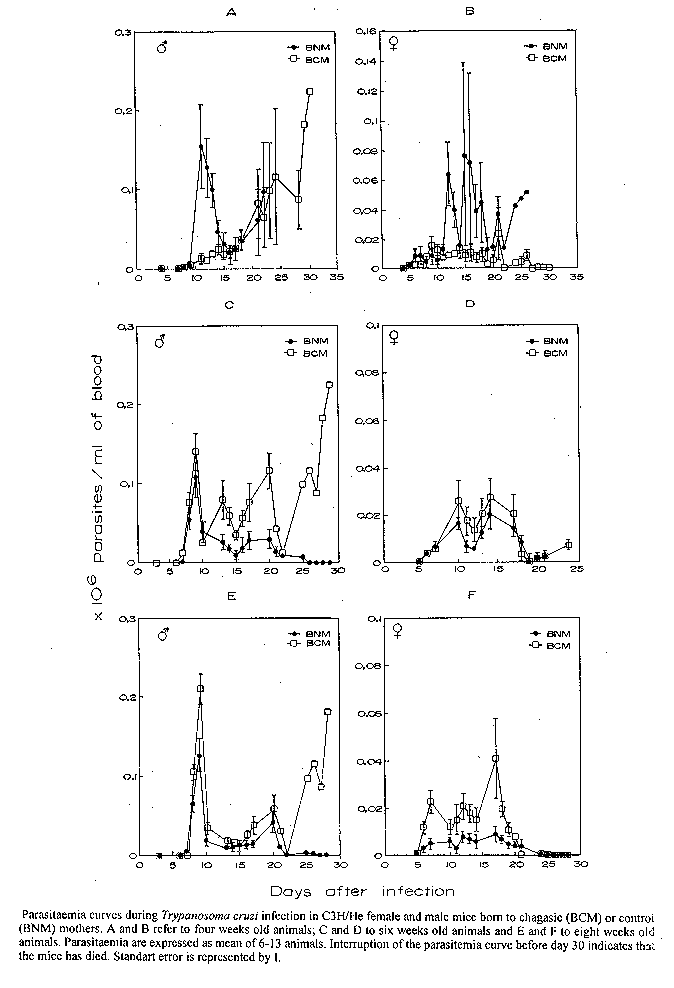
Offspring survival, birth rate and birth weigh - The ideal model of maternal-fetal versus T. cruzi infection was developed by infecting female mice groups via i.p. with differents inocula of a virulent and a non virulent clone of Y strain. Five hundred blood trypomastigotes of YP1 clone in C3H/He female mice were considered the ideal combination. Fertilization, pregnancy evolution and delivery were normally carried out. Three hundred and sixty seven and 346 births from 44 chagasic and 38 control mice mothers, respectively, were used to evaluate offspring survival, birth rate and birth weight. The mean of total birth per female was 8.40+/-4.73 to chagasic mothers compared to 9.11+/-3.77 of control mothers (p>0.05). The mean size of the litter obtained from chagasic mice mothers was 5.43+/-1.5 compared to 5.78+/-1.82 of the control mothers (p>0.05). The number of surviving and dead mice born to chagasic mice mothers were 175 (47.68%) and 192 (52.31%), respectively. These results were not significantly different from 180 surviving (52.02%) and 166 (47.98%) dead offsping born to control mice mothers (p>0.05). The weight of male mice born to chagasic mice mothers (13.06+/-1.76g) was significantly lower than that seen for male mice born to controls (16.52+/-1.27g) (p<0.001). On the other hand, there was no significant difference in the weight of females born to chagasic animals as compared to the ones born to controls (13.03g+/-2.04g versus 15.16g+/-1.71g, p>0.05). Congenital transmission evaluation - A total of 169 hemocultures were done, 71 for male and 98 for female. The congenital transmission rate was 1.18% (2/169). The positive hemocultures were observed in the female group. Evaluation of T. cruzi infection in offsprings - (a) Parasitaemia curves - Parasitaemia curves obtained after the infection of four, six and eight week old mice born to chagasic and control mothers are shown in the Fig. Four week old female mice born to chagasic mothers displayed lower parasitaemia levels than those born to controls (p<0.05) (Fig. 1B). Similar results were obtained with 4 week old male born to chagasic mothers up to the 15th day after infection (p<0.05) (Fig. 1A) . After this day the parasitaemia was not different for both male born to chagasic mothers and control ones. Six week old male mice born to chagasic mothers displayed higher parasitaemia curves than those from the control group (p<0.05) (Fig. 1C). In contrast, the parasitaemia curves displayed by six week old female mice born to chagasic mothers were not significantly different from those of the control group (p>0.05) (Fig. 1D). Eight week old female mice born to chagasic mothers clearly displayed higher parasitaemia curves than those observed in infected animals born to control group at the same age (p<0.05) (Fig. 1F). The parasitaemia curves displayed by eight week old male mice born to chagasic mothers were not significantly different from those obtained to eight week old male mice born to control mothers (p>0.05) (Fig. 1E). (b) Mortality rates (Table) - Mortality rates between four week old females born to chagasic mothers (50%) and their controls (90%) were significantly different (p<0.05). There were no significant differences in the mortality rates of four week old males born to chagasic mothers (83.3%) and those born to control mothers (83.3%) (p>0.05). Mortality rates observed in six week old females (62.5%) and males (81.8%) born to chagasic mother were not significantly different from those observed in female (66.7%) and male (69.2%) born to control mother with the same age (p>0.05). Although we observed significant differences in mortality rates between eight weeks females born to chagasic mothers (37.5%) and eight week females born to controls (0%) (p<0.05) this difference was not observed in the male group. The mortality rate for eight weeks old males born to chagasic mothers and for males born to controls were 85.7% and 62.5%, respectively (p>0.05). DISCUSSION
In this study important aspects of maternal-fetal relationship in Chagas' disease were demonstrated. Preliminary experiments had been performed in order to determine the ideal T. cruzi strain and inoculum size. As a result of the preliminary experiments, an ideal model of maternal-fetal versus T. cruzi infection was developed. As observed in natural conditions, mating, fertilization, pregnancy evolution and delivery were carried out successfully in our experimental model. Congenital transmission detected by hemoculture was 1.18% (2/169). In previous experiments we have shown that the positivity of this parasitological diagnostic technique was 93.3%. Other serological methods could produce doubtful results since circulanting antigens, immune complexes and offspring idiotypic sensitization with mother's antibodies have been described in Chagas' disease (Araujo 1976, Gottlieb 1981, Moreti et al. 1985, Gazzinelli et al. 1988). No significant differences could be observed in the litter size and in the total number of mice born to chagasic mothers versus control group. No significant differences were observed in mortality rates between offspring born to chagasic and those born to controls until four weeks of age, when the animals were weaned. However, four weeks old males born to chagasic mothers weighed significantly less than males born to controls. This effect has been reported previously in a model using BALB/c mice infected with the Tehuantepec strain for the study of maternal- fetal relationship (Carlier et al. 1987 a,b) and indicate that chronic infection has no influence on mouse reproductive capacity, but it can cause male abnormal growth. Our results clearly show that offspring born to chagasic mothers present decreased resistance to acquired T. cruzi infection after four weeks of age. This decreased resistance was expressed by higher levels of parasitaemia and higher mortality rates. Age was shown to be a crucial factor of this phenomenon. In murine T. cruzi infection, the maternal influence is a dynamic process: in four week old mice it has a protective effect, in six week old mice it has an unclear effect and in eight week old mice the decreased resistance is clearly demonstrated. Sex was also shown to be important. Although both sexes were probably exposed to the same maternal influence, females seem to control more effectively the decreased resistance. These findings are in agreement with classic literature data that showed that males are more susceptible to T. cruzi infection than females (Hauschka 1947, Rivera-Vanderpas et al. 1983). We have observed that even in four week old mice, when maternal anti- T. cruzi antibodies could still be detected at protective levels, males were less resistant. Four week old females born to chagasic mice mothers displayed lower levels of parasitaemia and lower mortality rates than the control group. On the other hand, four week old males only manage to control the parasitaemia up to the 15th day after infection. The decreased resistance is clear in eight weeks old animals. Eight week old females born to infected mothers displayed, both a high parasitaemia and a significantly higher mortality rate. This effect was not so evident in male mice, probably because of their high susceptibility to T. cruzi infection and also because of the high susceptibility of C3H/He mice strain itself (Trischmann et al. 1978). In fact it seems to be necessary to detect the decreased resistance an ideal combination of the inoculum size, the virulence of the parasite strain and the susceptibility of the mice strain to T. cruzi infection. Carlier et al. (1992) showed that in BALB/c males the phenomenon is more evident than in BALB/c female when 100 blood trypomastigotes/animal of Tehuantepec strain were used. In this last group the decreased resistance is more clearly shown when the inoculum is increased to 10.000 trypomastigotes/animal. Experiments with a highly virulent strain (100% mortality rate) could not demonstrate the phenomenon (unpublished data). These results are important as most T. cruzi strains isolated in nature have low virulence to mouse models in experimental infections (Schlemper Jr. 1982). The mechanism of such an interesting phenomenon is unknown. Many authors (Lewert & Mandlowitz 1969, Yamagushi et al. 1983, Watanabe et al. 1984, Haque et al. 1988, Dresser 1990, Koblasa et al. 1990, Suzuki & Kobayashi 1990, Telemo et al. 1991, Carlier et al. 1992), using different models, have suggested that antigens or maternal immune system products transferred by both placental and suckling routes could modulate the immune response of the offspring when their mothers had been challenged. Although further studies are still necessary to determine if this phenomenon also occurs in human Chagas' disease, we must keep in mind this possibility when working in endemic areas.
ACKNOWLEDGEMENTS
To Afonso da Costa Viana and Orlando Carlos Magno for diligent technical assistance and Drs Ricardo Moreira, Tereza Cristina RM de Oliveira and Lucia MC Galv o for the help in the preparation of the manuscript.
REFERENCES
Araujo FG 1976. Immunology of Chagas' disease. I. circulating antigens in mice experimentally infected with Trypanosoma cruzi. Rev Inst Med Trop S o Paulo 18: 443-439. Araujo SM, Chiari E 1988. Caracterizac o biologica de clones das cepas Y, CL e MR de Trypanosoma cruzi em camundongos C3H isogenicos. Mem Inst Oswaldo Cruz 83: 175-181. Azougue E, La Fuente C, Darras C 1985. Congenital Chagas' disease in Bolivia: epidemiological aspects and pathological findings. Trans R Soc Med Hyg 79: 176-180. Barousse AP, Eposto MO, Mandel S, Martinez FS 1978. Enfermedad de Chagas congenita en area no endemica. Medicina (Buenos Aires) 38: 611-615. Brener Z 1962. Therapeutic activity and criterion of cure in mice experimentally infected with Trypanosoma cruzi. Rev Inst Med Trop S o Paulo 4: 389-396. Camargo EP 1964. Growth and differentiation in Trypanosoma cruzi. I. origin of metacyclic trypanosomes in liquid media. Rev Inst Med Trop S o Paulo 6: 93-100. Carlier Y, Rivera MT, Truyens C, Goldman M, Lambert P, Flament J, Bauwens D, Vray B 1987a. Pregnancy and humoral immune response in mice chronically infected by Trypanosoma cruzi. Infect Immunol 55: 2496-2501. Carlier Y, Rivera MT, Truyens C, Puissant F, Milaire J 1987b. Interactions between chronic murine Trypanosoma cruzi infection and pregnancy: fetal growth retardation. Am J Trop Med Hyg 37: 534-540. Carlier Y, Rivera MT, Truyens C, Ontivero M, Flament J, Van Marck E, Maertelaer V 1992. Chagas' disease: decreased resistance to Trypanosoma cruzi acquired infection in offspring of infected mice. Am J Trop Med Hyg 46: 116-122. Cichero JA, Bonet AH, Santamaria N 1967. Estudio de la prevalencia de la enfermedad de Chagas en varones de 20 anos de edad en la Prov. de Santiago del Estero. Sem Med Argent 133: 1792. Corsini AC, Costa MG 1981. Immunossupression in mice infected with Trypanosoma cruzi. I. Evidence of polyclonal B cell activation in experimental infection mimicked by an extract prepared from circulating trypomastigotes. Rev Inst med Trop S o Paulo 23: 114-121. D'Imperio Lima MR, Joscowicz M, Coutinho A, Kipnis T, Eisen H 1985. Very large and isotipically atypical polyclonal plaque- forming cell responses in mice infected with Trypanosoma cruzi. Eur J Immunol 15: 201-203. Dresser OW 1990. The influence of maternal humoral responsiveness on the specific immunocompetence of the progeny in mice. J Reprod Immunl 18: 293-299. Espinal LF 1962. La enfermedad de Chagas congenita. Arch Venez Med Trop y Parasitol Med 4: 243-264. Gazzinelli RT, Morato MJF, Nunes RMB, Cancado JR, Brener Z, Gazzinelli G 1988. Idiotype stimulation of T lymphocytes from Trypanosoma cruzi-infected patients. J Immunol 140: 3167-3172. Gottlieb M 1981. A carbohydrate containing antigen from Trypanosoma cruzi and its detection in the circulation of infected mice. J Immunol 119: 465-470. Haque A, Cuna W, Pestel J, Capron A, Bonnel B 1988. Tolerance in rats by transplacental transfer of Dipetalonema vitae microfilarae: recognition of a putative tolerogen(s) by antibodies that inhibit antigen-specific lymphocyte proliferation. Eur J Immunol 18: 1167-1172. Hauschka TS 1947. Sex of host as a factor in Chagas' disease. J Parasitol 33: 399-404. Hoare CA 1972. The Trypanosomes of Mamals. Blackwell Scientific Publications, Oxford, 749pp. Koblasa F, Butler JE, Habe F 1990. Maternal-neonatal immunoregulation: suppression of de novo syntesis of IgG and IgA, but not IgM, in neonatal pigs by bovine colostrum, is lost upon storage. Am J Vet Res 51: 1407-1412. Lewert RM, Mandlowitz S 1969. Schistosomiasis: prenatal induction of tolerance to antigen. Nature 224: 1029-1031. Lugones A, Ledesma O, Jozanii LB 1979. Enfermedad de Chagas en la embarazada y Chagas congenita. Resumes de trabajos. II Simposio International Enfermedad de Chagas. FLAP: Buenos Aires. Minoprio PM, Eisen H, Forni L, D'Imperio Lima MR, Joskowicz M, Coutinho A 1986a. Polyclonal lymphocyte responses to murine Trypanosoma cruzi infection. I. Quantification of both T- and B-cell responses. Scand J Immunol 24: 661-668. Minoprio PM, Coutinho A, Joskowicz M, D'Imperio Lima MR, Eisen H 1986b. Polyclonal lymphocyte response to murine Trypanosoma cruzi infection. II. Cytotoxic T lymphocytes. Scand J Immunol 24: 669-679. Moretti E, Basso B, Vottero-Cima J 1985. Exoantigens of Trypanosoma cruzi. I. Conditions of their detection and immunogenic properties in experimentals infections. J Protozool 32: 150-153. Nisonoff A, Gurish MF, Kresna TF 1983. Fetal tranfer of a state of idiotipic suppression. Ann NY Acad Sci 418: 40-47. PAHO 1970. Report of a study group on Chagas' disease. PAHO Sci Publ 195. Passos EMC 1960. Molestia de Chagas na clinica obstetricia. Thesis. Faculdade de Medicina. Universidade do Rio de Janeiro, Brasil, 100pp. Rivera-Vanderpas MT, Rodriguez AM, Afchain D, Bazin H, Capron A 1983. Trypanosoma cruzi: variation in susceptibility of inbread rats. Act Tropica 40: 5-10. Schlemper Jr BR 1982. Caracterizac o de cepas do Trypanosoma cruzi isoladas de pacientes com diferentes formas clinicas da doenca de Chagas. PhD Thesis. Rio de Janeiro, Brasil, 131 pp. Suzuki Y, Kobayashi A 1990. Induction of tolerance to Toxoplasma gondii in newborn mice by maternal antibody. Parasitol Res 76: 424-427. Telemo E, Bailey M, Miller BG, Stokes CR, Bourne FJ 1991. Dietary antigen handling by mother and offspring. Scand J Immunol 34: 689-696. Trischmann TM, Tanowitz H, Wittner M, Bloom BR 1978. Trypanosoma cruzi: role of the immune response in the natural resistance of imbread strains of mice. Exp Parasitol 45: 160. Watanabe Y, Shimizu S, Yamaguchi N 1984. Effect of maternal antigenic stimulation on the active immune response of their offspring. Relationship between the immune reactivity of mother mice and the induction of suppression in their young. Scand J Immunol 20: 327-332. Wikler M, Demeur C, Dewasme G, Urbain J 1980. Immunoregulatory role of maternal idiotypes. Ontogeny of immune networks. J Exp Med 152: 1024-1035. Yamaguchi N, Shimizu S, Hara A, Saito T 1983. The effect of maternal antigenic stimulation upon the active immune responsiveness of their offspring. Immunology 50: 229- 238. This work was supported by grants from the Fundac o de Amparo a Pesquisa de Minas Gerais (FAPEMIG). SMA is fellowship from the Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq). ^+Corresponding author. Fax: 55-44-222.2754
Received 7 July 1995 FigureParasitaemia curves during Trypanosoma cruzi infection in C3H/He female and male mice born to chagasic (BCM) or control (BNM) mothers. A and B refer to four weeks old animals; C and D to six weeks old animals and E and F to eight weeks old animals. Parasitaemia are expressed as mean of 6-13 animals. Interruption of the parasitemia curve before day 30 indicates that the mice has died. Standart error is represented by I. TABLE Mortality rates during Trypanosoma cruzi infection in C3H/He female and male mice born to chagasic (BCM) or to normal mother (BNM)
Age at the Male Female time of BCM BNM BCM BNM infection------------------------ -------------------------- (weeks) no./N^a % no/N % no/N % no/N % --------------------------------------------------------------- 4 5/6 83.3 5/6 83.3 6/12 50^b 9/10 90 6 9/11 81.1 9/13 69.2 5/8 62.5 4/6 66.7 8 6/7 85.7 5/8 62.5 3/8 37.5^b 0/9 0 a: number of dead mice/total of animals; b: p<0.05 Copyright 1996 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc96042a.gif] |
| |||||||||

{kind=link}
{kind=link}