 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Aspectos Bionomicos de Squamatoides trivittatus (Diptera, Sarcophagidae) sob Condic es de Laboratorio
Ricardo JB Salviano, Rubens Pinto de Mello*, Lilian CNH Beck, Jose Mario d'Almeida
Laboratorio de Biologia e Controle de Insetos Vetores, Departamento de Biologia *Laboratorio de Diptera, Departamento de Entomologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045- 900 Rio de Janeiro, RJ, Brasil
Code Number: OC96049
Size of Files:
Text: 24.5K
Graphics: Line Drawings (gif) - 12.5K[TABLES AND FIGURES AT END OF TEXT] Bionomical aspects of de Squamatoides trivittatus (Diptera, Sarcophagidae) under laboratory conditions - Experiments with Squamatoides trivitattus were carried out in two different controlled temperatures (16+/-1 C/50-60% RH and 27+/-1 C/70-80% RH). The viability of larvae and pupae at 27 C was 89.82% and 92.75% respectively. Larvae did not develop at 16 C. Larval development lasted for 20+/-4 hr, 16+/-8 hr and 60+/-7 hr for the first, second and third instars, respectively, completing a total of 96+/-6 hr. The mean pupal period lasted for 15.7+/-1.6 days. In longevity tables for the adults, life- Expectancy for 50% of the colony submitted to 16 C was of 1.78 weeks for males and 2.42 for females. At 27 C a life-Expectancy of 1.15 weeks for males and 0.78 week for females was recorded. The average life-spans for males and females at 16 C were 3.5+/- 2.0 and 3.8+/-2.6 weeks, respectively, and 1.9+/-1.2 weeks for both sExes. At 27 C, the longevity recorded was of 2.1+/-1.3 weeks for males and 1.7+/-1.1 week for females.
O genero Squamatoides Curran, 1927 esta representado por tres especies: S. trivittatus Curran, 1927, S. spitzi (Lopes 1933) e S. stahli Dodge, 1966, sendo que apenas as duas primeiras s o encontradas no Brasil. S. trivittatus esta restrita ao Campo Cerrado da America do Sul; no Brasil e encontrada nos estados de Mato Grosso, Goias, Minas Gerais e S o Paulo (Lopes 1969). Linhares (1981), estudando dipteros muscoides, em S o Paulo, verificou que S. trivittatus apresenta um elevado grau de sinantropia. Em areas que apresentam condic es ambientais muito proximas a estudada por Linhares, Mendes (1991) obteve 33,33% dos especimens coletados em fezes humanas, o que confere importancia a este Sarcophagidae como potencial vetor de agentes enteropatogenos. Apesar do interesse medico e sanitario dos dipteros desta familia, s o escassos os estudos sobre sua biologia. Em sua maioria, os trabalhos relacionados com as fases imaturas avaliam o periodo de desenvolvimento, a viabilidade e a competic o larval em diversas condic es de temperatura e umidade relativa (Kamal 1958, House 1977, Levot et al. 1979, Baxter & Morrison 1983; Jiron & Bolanos 1986, Ferraz 1992a,b). Os artigos relacionados a fase adulta est o em grande parte relacionados aos aspectos comportamentais (preferencia de substrato para postura e alimentac o), biogeograficos (sinantropia), longevidade e capacidade reprodutiva (Kamal 1958, Ferreira 1978, Linhares 1979, 1981, d'Almeida 1983, 1984, 1986, 1988, 1989, 1993, Ferraz 1992a,b). Tais aspectos, dentre outros, podem ser analisados atraves de tabelas de vida, que nos permitem avaliar satisfatoriamente todo o processo de vida dos insetos e relaciona-lo a diversos fatores, como temperatura, dieta, efeito de grupo (Pearl & Parker 1921, Pearl et al. 1927, Jackson 1936, Mendes 1961, Carey 1987, Bressan & Teles 1991). Devido a inExistencia de trabalhos sobre a biologia de S. trivittatus, foram avaliados alguns aspectos de suas fases evolutivas: periodo e viabilidade larval e pupal, raz o sExual, longevidade, curva de sobrevivencia de adultos em diferentes temperaturas e umidades. Foram elaboradas tabelas de Expectativa de vida.
MATERIAIS E METODOS
Os Experimentos foram realizados em camaras climatizadas reguladas a 16+/-1 C e 27+/-1 C de temperatura e 50-60% e 70-80% de umidade relativa do ar (URA). Essas temperaturas foram escolhidas por corresponderem as medias das maximas e minimas durante 27 anos, no Distrito Federal. A colonia foi estabelecida a partir de insetos coletados em areas de cerrado e urbana da cidade satelite Taguatinga (DF) no periodo de 27 de marco a 04 de abril de 1992. Para observac o da viabilidade larval, utilizaram-se larvas de colonias mantidas em temperatura ambiente (25-27 C). Setenta e oito grupos de 10 larvas de primeiro estadio foram introduzidos em recipientes plasticos (5 cm altura x 6 cm diametro), contendo, cada um, 20g de sardinha com 24h de decomposic o (2g/larva). Desses grupos, 20 foram levados para camara climatizada a 16 C e os demais mantidos em camara a 27 C. As observac es diarias foram feitas em dois horarios: 8h e 17h30 min. Para avaliar a durac o do periodo larval foram utilizadas 720 larvas obtidas de larviposturas de uma colonia de individuos da sExta gerac o. Foram separados grupos de 20 larvas em 36 amostras. A cada duas horas uma amostra era retirada da camara climatizada (27 C e 70-80%) e as larvas eram sacrificadas e fixadas em San Jean (600 ml de agua destilada, 300 ml de alcool a 95%, 100 ml de formalina e 10 ml de acido acetico) a quente, e conservadas em alcool a 70%, para posterior verificac o do respectivo estadio. Quanto a fase pupal (durac o e viabilidade) e raz o sExual, foram usadas 1264 larvas maduras recem-saidas dos substratos de criac o. Apos pesadas e individualizadas, 200 foram transferidas para a camara climatizada a 16 C e as demais para camara a 27 C, registrando-se diariamente as que puparam, as pupas mortas e a emergencia dos imagos. Apos emergencia e sExagem, 14 amostras contendo 15 casais cada uma, foram separadas em dois grupos: o primeiro, contendo nove amostras, submetido a 27 C e o segundo, com cinco amostras, a 16 C. Como fonte diaria de proteina e substrato de larviposic o, utilizou-se sardinha, retirada do congelador, e mantida por 24h em temperatura ambiente, fornecida, para cada gaiola, na proporc o de 10 g por pote; alem de agua ad libitum e acucar mascavo. Registros diarios do numero de mortos em cada amostra foram efetuados com o objetivo de se construir as tabelas de vida, as curvas de sobrevivencia e a estimativa das longevidades medias. As longevidades para os dois tratamentos foram transformadas em [Square Root of x] e submetidas a analise das variancias. As medias foram comparadas pelo teste "t" ao nivel de significancia de 5%. As tabelas de vida foram baseadas em Carey (1987) e Silveira Neto et al. (1976), que sugerem iniciar as mesmas com a projec o para 1000 Exemplares, que e sua raiz. Os parametros est o organizados em colunas, em func o do intervalo de idade (x), em semanas. Lx representa o numero de sobreviventes no comeco da idade x; dx e a frac o de insetos mortos entre as idades x e x+1, estimada por L[x] - L[x+1]. Ex representa a estrutura etaria, calculado por L[x] + L[x+1] / 2. O valor de Tx obtem-se pelo somatorio de Ex + E[x+1] + E[x+2] + ... + E[w], onde w e o ultimo estagio. A esperanca de vida (Ex) foi estimada por T^x / L[x]. 1000qx e a coluna de risco, estimada por 1000 d^x / L[x].
RESULTADOS E DISCUSS O
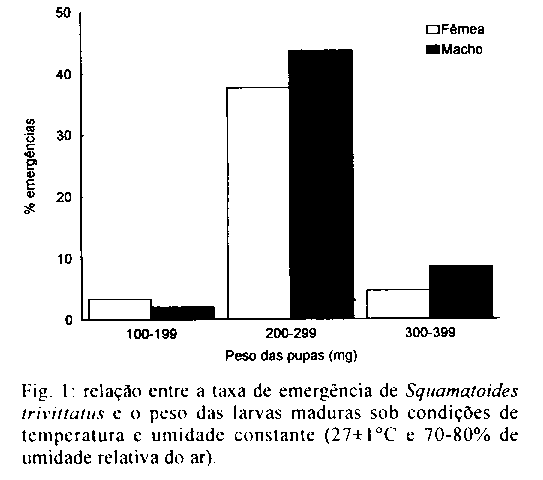
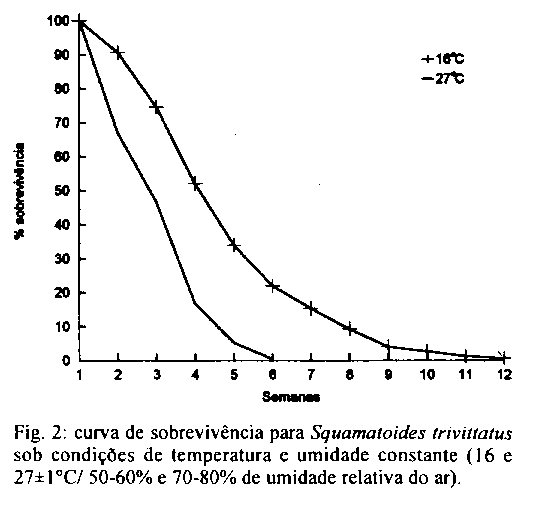
As larvas submetidas a 16 C so se desenvolveram ate o segundo estadio. Tal fato pode ter ocorrido devido a Exposic o prolongada a temperatura e umidade baixas, provocando um ressecamento do substrato e, consequentemente, a morte das larvas; o mesmo foi observado durante a avaliac o do periodo pupal, quando as 200 larvas maduras chegaram a pupar, mas n o ocorreu emergencia. Apos tres meses de observac o, admitiu-se a hipotese de as pupas terem entrado em diapausa. Para esta avaliac o, as pupas foram divididas em tres grupos. No primeiro, atraves de dissecac o avaliou-se o estado dos imagos, verificando a sua total desidratac o. O segundo foi Exposto a temperatura de 25 C e 60- 70% de URA e o terceiro a 27 C e 80-90% de URA, e em ambos verificou-se que as larvas maduras chegaram a pupar, porem n o emergiram devido a total desidratac o do imago. Em algumas especies, a temperatura baixa n o provoca a completa inviabilidade das fases imaturas, como em Parasarcophaga crassipalpis e Boettcherisca peregrina, que apresentam um periodo pupal minimo de 56 e 50 dias, respectivamente, quando submetidos a temperatura de 15 C (Nishida 1984). A 27 C a viabilidade media larval foi de 89,82%, valor proximo aos obtidos por Madubunyl (1986) para Sarcophaga haemorrhoidalis (80,69%, a 23-28 C). Resultados semelhantes tambem foram verificados para Peckia chrysostoma e Adiscochaeta ingens, submetidos a temperatura de 25,9 C e 79% de URA, com 97% e 93% de sobrevida, respectivamente (Ferraz 1992a). O periodo larval para S. trivittatus a 27 C durou 96+/-6 h, onde o primeiro estadio teve durac o de 20+/-4 h, o segundo, de 16+/-8 h e o terceiro de 60+/-7 h. O periodo larval do Sarcophagidae, P. chrysostoma, analisado por Ferraz (1992a), foi de 89 h a 25,9 C e 95 h a 27 C, bem proximo ao obtido para S. trivittatus. Levot (1979) verificou que, a 27-28 C, larvas de P. crassipalpis alcancaram o final do terceiro estadio apos 69 h, com peso maximo e minimo de 235,7 mg e 173,8 mg, respectivamente. Esta mesma especie foi estudada por Nishida (1984), onde o periodo e peso maximo obtido em 25 C foi de oito dias e 152,6 mg; a 30 C chegou a sete dias com 135,9 mg. Para B. peregrina, esse autor obteve os seguintes pesos maximos e durac es larvarias nas temperaturas de 25 C e 30 C: seis dias com 115,8 mg, e cinco dias com 87 mg, respectivamente. A viabilidade pupal para S. trivittatus foi de 92,75%. Kamal (1958) obteve resultados semelhantes para Sarcophaga spp. Madubunyl (1986), estudando Bercaea haemorrhoidalis, observou que a viabilidade pupal foi de 89,83% a 23-28 C. De acordo com Ferraz (1992a) a 25,9 C e 79% de URA a viabilidade pupal para P. chrysostoma foi de 69,9% e para A. ingens 92,8%. O periodo pupal medio foi de 15,7+/-1,6 dias, com o pico de emergencia no 15^o dia, para 37% das 1064 pupas. Dos adultos emergidos, 54,36% foram machos e 45,64% femeas; a emergencia de adultos foi maior nas pupas que pesavam entre 200 e 299 mg (Fig. 1). Os pesos maximos e minimos das larvas que originaram femeas e machos foram de 320-120 mg e 370-150 mg, respectivamente. O peso medio para os machos foi de 257+/-33 mg e 238+/-34 mg, para as femeas. A raz o sexual foi de 0,6 nos imagos provenientes de larvas de 100-199 mg, 1,2 para as larvas de 200-299 mg e 1,8 para as larvas de 300-399 mg. Segundo Ferraz (1992a), o periodo pupal de P. chrysostoma e A. igens a 18 C foi 23,5+/-1,5 dias e 33+/-3 dias, respectivamente. O periodo pupal dos Sarcophagidae, em temperaturas proximas a 27 C geralmente fica na faixa de 10-20 dias (Nishida 1984, Jiron & Bolanos 1986, Ferraz 1992a), o que concorda com os presentes resultados. Na Tabela I s o apresentadas as Expectativas de vida para machos e femeas de S. trivittatus, nas temperaturas e umidades relativas propostas. A Expectativa de vida para 50% da colonia submetida a 16+/-1 C e 50-60% de URA foi de 1,78 semanas para os machos e 2,42 semanas para as femeas. Ja a colonia mantida a 27+/-1 C e 70-80% de URA foi de 1,15 semana para os machos e 0,78 semana para as femeas. Estes valores juntamente com os apresentados na Tabela II, deixam claro que a baixa temperatura, a femea de S. trivittatus apresenta uma longevidade maior do que as do macho, provavelmente devido a sua baixa atividade metabolica. A 27 C ocorreu o inverso onde as femeas apresentaram uma longevidade menor do que os machos, devido ao maior desgaste (aumento do metabolismo). Um dos aspectos que podem ter evidenciado este fato e o desgaste devido ao desenvolvimento ovariano, como tambem, pode-se sugerir que esta constatac o esteja ligada a algumas caracteristicas peculiares do ecossistema onde S. trivitattus ocorre. Estes resultados n o corroboram com Mackerras (1933) que estudou femeas de varias especies de moscas que vivem mais do que os machos. Ferraz (1992a) tambem obteve maior longevidade para femeas de P. chrysostoma e A. ingens . A Fig. 2 apresenta as curvas de sobrevivencia que tendem para o tipo III (Silveira Neto 1976, Carey 1987), onde dx decresce gradativamente a cada intervalo x (Tabela I). Analisando as curvas, pode-se verificar que a sobrevivencia foi maior a 16 C, onde dx aumentou rapidamente ate a 4^a semana, decaindo mais lentamente ate a 7^a, quando aumentou novamente ate a ultima semana. A 27 C, ocorreu o mesmo para dx, sendo que terminam na 7^a semana. Os sarcofagideos de regi es tropicais, apresentam diferencas biologicas quando est o em seus respectivos ambientes naturais, onde os fatores abioticos modificam-se constantemente ao longo do dia, e as suas necessidades alimentares n o s o limitadas ou Excluidas, ao contrario do que ocorre quando criados em laboratorio. No caso de S. trivittatus, acredita-se que as condic es do Cerrado, onde ela e endemica, influam marcadamente na sua bionomia.
AGRADECIMENTOS
Ao Dr. Hugo de Souza Lopes, in memorian, pela iniciac o nos estudo dos dipteros caliptrados, pelo apoio, incentivo e Exemplo profissional. A Vanderleia Cristina de Oliveira, pela ajuda na manutenc o das colonias. A Dra. Marli Maria Lima pela revis o e sugest es no tExto.
REFER NCIAS
Baxter JA, Morrison PE 1983. Dynamics of growth modified by larval population in the flesh fly, Sarcophaga bullata. Can J Zool 61: 512-517. Bressan S, Teles MC 1991. Longevidade e curva de sobrevivencia de tres especies do genero Anastrepha Schiner, 1868 (Diptera, Tephritidae) em laboratorio. Rev bras Ent 35: 685-690. Carey J 1987. Demographic methods in fruit fly research: survival, reproduction and mass-rearing. Ed. Global Fruit Working Group, California, 64pp. d'Almeida JM 1983. Sinantropia de dipteros caliptrados na area metropolitana do Rio de Janeiro, Tese de Mestrado, UFRRJ, Rio de Janeiro, 193pp. d'Almeida JM 1984. Sinantropia de Sarcophagidae (Diptera) na regi o metropolitana do Estado do Rio de Janeiro. Arq Univ Fed Rur Rio J 7: 101-110. d'Almeida JM 1986. Substratos utilizados para a criac o de dipteros caliptrados em uma area rural do Estado do Rio de Janeiro. Arq Univ Fed Rur Rio J 9: 13-22. d'Almeida JM 1988. Substratos utilizados para a criac o de dipteros caliptrados em uma area urbana do Municipio do Rio de Janeiro. Mem Inst Oswaldo Cruz 83: 201-206. d'Almeida JM 1989. Substrato utilizados para a criac o de dipteros caliptrados no Jardim Zoologico do Rio de Janeiro (RIO- ZOO). Mem Inst Oswaldo Cruz 84: 257-264. d'Almeida JM 1993. Capture of calliptrate flies with different breeding substrates on beaches in Rio de Janeiro, RJ, Brazil. Mem Inst Oswaldo Cruz 88: 215-220. Ferraz MV 1992a. Estudos comparados do desenvolvimento biologico e do comportamento de Peckia chrysostoma (Wiedemann, 1830) e Adiscochaeta ingens (Walker, 1849) (Diptera: Sarcophagidae) em condic es de laboratorio. Tese de Mestrado, Instituto Oswaldo Cruz, Rio de Janeiro, 138pp. Ferraz MV 1992b. Comparison of the reproductive between isolated Peckia chrysostoma (Wiedemann, 1930) and Adiscochaeta ingens (Walker, 1849) (Diptera: Sarcophagidae) females reared in laboratory. Mem Inst Oswaldo Cruz 87: 131-139. Ferreira MJM 1978. Sinantropia de dipteros muscoides de Curitiba, Parana, I. Calliphoridae. Rev Bras Biol 38: 445-454. House HL 1977. Interaction between amino acids and glucose in larval nutrition of the fly Agria housei (Diptera: Sarcophagidae) at low temperature. Can Ent 109: 87- 91. Jackson CHN 1936. Some new methods in the study of Glossina morsitans. Proc Zool Soc Lond 193: 811-896. Jiron LF, Bolanos R 1986. Biology and larval morphology by scannig electron microscopy of Panttonella intermutans Walker (Diptera, Sarcophagidae). Rev bras Ent 30: 27- 30. Kamal AS 1958. Comparative Study of Thirteen Species of Sarcosaprophagous Calliphoridae and Sarcophagidae (Diptera) I. Bionomic. Ann Ent Soc Am 51: 261-271. Levot GW, Brown KR, Shipp E 1979. Larval growth of some calliphorid and sarcophagid Diptera. Bull Ent Res 69: 469- 475. Linhares AX 1979. Sinantropia de dipteros muscoides de Campinas. Tese de Mestrado. Universidade Estadual de Campinas, Campinas, SP, 129 p. Linhares AX 1981. Synanthropy of Calliphoridae and Sarcophagidae (Diptera) in the city of Campinas, S o Paulo, Brazil. Rev bras Ent 25: 189-215. Lopes HS 1969. Family Sarcophagidae. In A Catalogue of the Diptera of the Americas South of the United States. Museu de Zoologia da Universidade de S o Paulo. Fasc. 103, 88 pp. Mackerras MJ 1933. Observations on the life-histories, nutrutional requirements and fecundity of blowflies. Bull Entomol Res 24: 353-562. Madubunyl LC 1986. Laboratory life history parameters of the red- tailed fleshfly, Sarcophaga haemorrhoidalis (Fallen) (Diptera: Sarcophagidae). Insect Sci Appl 7: 617-621. Mendes J 1991. Relac o entre atratividade por iscas e estagios de desenvolvimento ovariano em femeas de dipteros muscoides sinantropicos de Campinas, SP. Tese de Mestrado, Universidade Estadual de Campinas, Campinas, SP, 129 pp. Mendes LOT 1961. Longevidade de Dysdercus. III. Machos Adultos de Dysdercus mendesi Bloete em condic es de laboratorio. Bragantia 20: 471-494. Nishida K 1984. Experimental studies on estimation of postmortem interval by means of fly larvae infesting human cadavers. Nippon-Hoigaku-Zasshi 38: 24-41. Pearl R, Parker SL 1921. Experimental studies on the duration of life. I. Introductory discussion of the duration of life in Drosophila. Amer Nat 55: 481-509. Pearl R, Miner JR, Parker SL 1927. Experimental studies on the duration of life. XI. Density of population and life duration in Drosophila. Amer Nat 61: 289-318. Silveira Neto S, Nakano O, Barbin D, Nova NAV 1976. Manual de ecologia dos insetos. Agronomica Ceres, S o Paulo, p. 243- 245.
Recebido em 21 de julho de 1995 Fig. 1: relac o entre a taxa de emergencia de Squamatoides trivittatus e o peso das larvas maduras sob condic es de temperatura e umidade constante (27+/-1 C e 70-80% de umidade relativa do ar). Fig. 2: curva de sobrevivencia para Squamatoides trivittatus sob condic es de temperatura e umidade constante (16 e 27+/-1 C/ 50-60% e 70-80% de umidade relativa do ar). TABELA I Expectativa de vida de machos e femeas de Squamatoides trivittatus a 16 e 27+/-1 C/ 50-60% e 70-80% de umidade relativa do ar
Macho
x Lx dx Ex
16 C 27 C 16 C 27 C 16 C 27 C
-------------------------------------------------
1 1000 1000 120 285 940,0 857,5
2 880 715 93 228 833,5 601,0
3 787 487 280 244 647,0 365,0
4 507 243 160 169 427,0 158,5
5 347 74 120 74 287,0 37,0
6 227 - 54 - 4,0 -
7 173 - 96 - 125,0 -
8 77 - 58 - 48,0 -
9 19 - 19 - 9,5 -
10 - - - - - -
11 - - - - - -
12 - - - - - -
Macho continued/
x Tx Ex 100qx
16 C 27 C 16 C 27 C 16 C 27 C
----------------------------------------------------
1 3321,0 2019,0 3,32 2,02 12,0 28,5
2 2381,0 1161,5 2,71 1,63 10,6 31,9
3 1547,5 560,5 1,97 1,15 35,6 50,0
4 900,5 195,5 1,78 0,81 31,6 69,7
5 473,5 37,0 1,37 0,50 34,6 100,0
6 186,5 - 0,82 - 47,1 -
7 182,5 - 1,06 - 55,6 -
8 57,5 - 0,75 - 75,0 -
9 9,5 - 0,50 - 100,0 -
10 - - - - - -
11 - - - - - -
12 - - - - - -
Femea
x Lx dx Ex
----------------------------------------------------
16 C 27 C 16 C 27 C 16 C 27 C
----------------------------------------------------
1 1000 1000 67 378 966,5 811,0
2 933 622 227 178 819,5 533,0
3 706 444 173 355 619,5 266,5
4 533 89 200 59 433,0 59,5
5 333 30 120 23 273,0 18,5
6 213 7 27 7 199,5 3,5
7 186 - 53 - 159,5 -
8 133 - 67 - 99,5 -
9 66 - 13 - 59,5 -
10 53 - 27 - 39,5 -
11 26 - 13 - 19,5 -
12 13 - 13 - 6,5 -
Femea continued/
x Tx Ex 100qx
---------------------------------------------------
16 C 27 C 16 C 27 C 16 C 27 C
---------------------------------------------------
1 3695,0 1692,0 3,70 1,69 6,7 37,8
2 2728,5 881,0 2,92 1,42 24,3 28,6
3 1909,0 348,0 2,70 0,78 24,5 80,0
4 1289,5 81,5 2,42 0,92 37,5 66,7
5 856,5 22,0 2,57 0,73 36,0 75,0
6 583,5 3,5 2,74 0,50 12,5 100,0
7 384,0 - 2,07 - 28,6 -
8 224,5 - 1,69 - 50,0 -
9 125,0 - 1,89 - 20,0 -
10 65,5 - 1,24 - 50,0 -
11 26,0 - 1,00 - 50,0 -
12 6,5 - 0,50 - 100,0 -
X: Intervalo De Idade Em Unidades De Tempo (Semanas); TABELA II Longevidade media para machos e femeas de Squamatoides trivittatus sob condic es de temperatura e umidade constante
(16 e 27+/-1 C/ 50-60% e 70-80% umidade relativa do ar)
Longevidade media (semanas)
----------------------------------
Temperatura Macho Femea Macho e femea
-------------------------------------------------
16 C 3,5+/-2,0a 3,8+/-2,6a 1,9+/-1,2
(0,4-8,7) (0,4-10,8)
27 C 2,1+/-1,3b 1,7+/-1,1b 3,7+/-0,7
(0,3-4,8) (0,3-5,8)Medias Seguidas Pelas Mesmas Letras N o Diferem Entre Si Pelo Teste "T" Ao Nivel De 5%; Longevidade Minima E Maxima Em Semanas. Copyright 1996 Fundacao Oswaldo Cruz
The following images related to this document are available:Line drawing images[oc96049b.gif] [oc96049a.gif] |
| |||||||||

{kind=link}
{kind=link}