 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Biology of Malarial Parasite in the Mosquito - A Review
Amauri Braga Simonetti
Departamento de Microbiologia, Instituto de Biociencias,
Universidade Federal do Rio Grande do Sul, Rua Sarmento Leite
500, 90050-170 Porto Alegre, RS, Brasil
Received 20 December 1995
Code Number: OC96099
Sizes of Files:
Text: 126K
Graphics: line drawings (gif) - 15.4K
[TABLES AND FIGURES AT END OF TEXT] The purpose of this review is to summarize the biology of Plasmodium in the mosquito including recent data to contribute to better understanding of the developmental interaction between mosquito and malarial parasite. The entire sporogonic cycle is discussed taking into consideration different parasite/vector interactions and factors affecting parasite development to the mosquito.
Key words: malaria - mosquito - sporogonic cycle - cell biology The control and prevention of malaria has been pursued for a long time. Although campaigns against malaria were initially successful in some areas, the emergence of resistance of the parasite to drugs and of the mosquito vector to insecticides, combined with the difficulties in implementing and maintaining effective control schemes have led to a resurgence of the disease in many parts of the world (Wernsdorfer 1991, Schapira et al. 1993, Roush 1993). Much of the current work on malaria focuses on immunological aspects of the disease and there has been remarkable progress in the identification of a variety of parasite antigens in different stages of the parasite development, some of which may be included in a future vaccine (Nussenzweig & Nussenzweig 1989, Mitchel 1989, Targett 1989, Nussenzweig 1990, Hommel 1991, Good 1991a). However, experimental and field studies have shown the complexity of the immune response to parasites, indicating that an efficient malaria vaccine is difficult to achieve (Cattani 1989, Good 1991b, 1992, Philips 1992). Therefore, for control of the disease greater understanding of the biological mechanisms involved in host/parasite/vector interactions is essential. In recent years an increasing number of studies have concentrated on the mosquito stages of the parasite development. Comprehensive reviews on biological, ultrastructural, biochemical, molecular and immunological aspects of the parasite and the vector can be obtained in the literature (Sinden 1978, 1983, 1984, Carter & Graves 1988, Carter et al. 1988, Alano & Carter 1990, Billingsley 1990, Crampton et al. 1990, 1994, Alano 1991, Brey 1991, Coluzzi 1992, Kaslow et al. 1994a, Sinden et al. 1996). This review foccuses on the cell biology of malarial parasites at different stages of its development in the mosquito and also includes factors that may affect the parasite infectivity. The sporogonic cycle Some erythrocytic parasites differentiate into sexual forms called gametocytes. Mature and functional gametocytes ingested by an appropriate species of mosquito in a bloodmeal are stimulated to transform into the stages which establish the parasites in their vector (Garnham 1966). Under the influence of changes in the mosquito midgut environment the gametocytes become extracellular within 8-15 min of ingestion. After emergence from the red blood cell (exflagellation) the male gametes fertilize the female gametes within 60 min of ingestion of blood. The fertilized macrogamete (zygote) differentiates into a single motile ookinete over the next 10- 25 hr, which migrates from the bloodmeal through the midgut wall to form an oocyst underneath the basal lamina of the midgut. Each oocyst produces many thousands of invasive sporozoites over a period of 7-12 days. The sporozoites escape from the oocyst and then invade the salivary glands, here they stay for possibly very long periods until injected into another vertebrate host when the next bloodmeal is taken (Sinden 1984, Carter & Graves 1988). A diagram of the sporogonic cycle in the mosquito is shown in Figure. Development of gametocytes (gametocytogenesis) As with other members of the order Haemosporina, gametocytes of Plasmodium can arise from merozoites from pre- erythrocytic parasites. Nevertheless, during natural infections, most gametocytes arise from merozoites of blood- stage origin (Alano & Carter 1990). Two alternative possibilities by which malaria parasites could become committed to either sexual or asexual development have been proposed (Inselburg 1983, Mons 1985, 1986, Bruce et al. 1990): first, the merozoite is not committed at the time of invasion of a red blood cell. During early growth (as a ring form), the parasite is suceptible to factors that will commit it to either sexual or asexual development. Second, during growth of an asexual parasite, environmental factors influence it so that at maturity the schizont produces merozoites that are precommitted to form asexual parasites or committed to form gametocytes upon subsequent invasion of a red blood cell. A number of studies have attempted to identify factors that may influence the differentiation and production of sexual stage parasites including cAMP, presence of antimalarial drugs and other environmental factors associated to the host such as the presence of serum undetermined components (reviewed by Sinden 1983, Mons 1986, Carter & Graves 1988, Dearsly et al. 1990). In a recent report (Schneweis et al. 1991) it was claimed that the production of infective gametocytes in vitro can be enhanced by products of haemolysis but the nature of the erythrocytic factor was not identified. Gametocyte development takes longer than asexual schizogony, e.g. 26 hr as opposed to 22.5 hr in P. berghei (Mons et al. 1985) or in the more extreme case of P. falciparum 8-12 days as opposed to 48 hr (Sinden 1983). The proportion of parasites that develop into gametocytes varies greatly during the course of natural infections even at its peak but is very low in relation to the total parasitaemia (Smalley et al. 1981). The growth and differentiation of gametocytes of P. falciparum has been divided into five stages (I to V) covering about eight days from merozoite invasion to mature gametocyte, each stage being distinguished by successive changes in the organization of the cell (Hawking et al. 1971). Little is known about commitment of gametocytes to be male or female. The fact that both female and male can be produced by haploid blood-stage parasite, the sex of a gametocyte does not result from chromosomal differences between both types of cell but their development must be due to selective gene expression. In general, the number of female gametocytes predominates over the number of male, but this predominance may vary at different times between cloned infections (Cornelissen 1988, Schall 1989). Gametogenesis Mature and functional gametocytes ingested by the mosquito in a bloodmeal are stimulated by the midgut environment to transform into gametes. Studies indicate that various triggers induce gametocytes to undergo differentiation. Microgameto- genesis in vitro is, optimally, dependent upon a rise in pH (Nijhout & Carter 1978), a rise in pCO2 and bicarbonate levels (Carter & Nijhout 1977, Nijhout & Carter 1978), a fall in temperature of a few degrees below that of the vertebrate host (Sinden & Croll, 1975) or a very potent factor termed mosquito exflagellation factor (MEF). The latter is a small heat stable molecule from the mosquito head and gut which stimulates gametogenesis via a bicarbonate- and pH-independent mechanism (Nijhout 1979). Recently, Kawamoto et al. (1991) showed in vitro that induction of exflagellation of P. berghei is triggered by a rise in the intracellular pH (pHi) which is mediated by Ca^++ and cGMP regulation. pHi can be modulated by alkaline media and is controlled by a complex series of interdependent ion pumps and channels controlling Na^+, K^+, Cl^- and HCO3^- transport between the parasite and the environment. Other influential factors described include cAMP analogues and inhibitors of phosphodiesterase (Martin et al. 1978). The duration of microgametogenesis is both temperature and species dependent, e.g. at 20-22 C it may take 7-15 min for P. falciparum in vitro, although exflagellation may be detected after shorter periods in the fluid excreted by feeding Anopheles (Sinden 1983). There is no evidence that exflagellation is influenced by factors released by digestion of the blood meal since digestion normally begins several hours later (Graf et al. 1986). Microgamete formation involves three mitotic divisions with a rapid assembly of eight axonemes on the single microtubule organizing centre that divides and passes to the spindle poles. This division simultaneously segregates the genome and the axoneme so that each of the eight emergent gametes receives a single axoneme and haploid genome, both being connected to a common microtubule organizing centre. After exflagellation the microgametes, normally bearing a single axoneme, a single condensed nucleus and a single kinetosome with its sphere and granule at the distal end, are torn from the microgametocyte surface and rapidly move away into the blood meal (Sinden & Croll 1975). Macrogametogenesis at the morphological level involves little more than escape from the host cell (Sinden 1984). At the cellular level there is de novo synthesis of the proteins which are expressed on the surface of macrogamete (Kumar & Carter 1984). It was recently identified a gametocyte specific protein of P. falciparum called Pf11-1 and there is some evidence that this protein might be involved in the emergence of gametes from the infected erythrocyte (Scherf et al. 1992, 1993). Interactions between the vertebrate host and the parasite do not cease when the blood meal is taken. Following induction of gametogenesis the parasite is liberated into the blood meal. It has been shown that the gametes are susceptible to the phagocytes in the host's blood (Rutledge et al. 1969, Sinden & Smaley 1976) and that they are susceptible to activation of the components of the complement pathway (Grotendorst et al. 1986, 1987). Zygote formation (fertilization) and ookinete development Fertilization is rapid usually occurring within 1 hr of gamete formation, the plasmalemma of the two gametes fuse and the microgamete axoneme and nucleus enter the macrogamete cytoplasm (Sinden 1983). During zygote development, structural changes occur and its transformation to ookinete is in part determined by the gradual assembly of the apical complex including the collar and pollar rings, rhoptries and micronemes (Canning & Sinden 1973, Davies 1974, Sinden 1978). Intensive protein synthesis also begins in the fertilized macrogamete and continues in the zygote and developing ookinete as reported for different Plasmodium species (Kaushal et al. 1983a, Kaushal & Carter 1984, Kumar & Carter 1985, Vermeulen et al. 1985a, 1986, Sinden et al. 1987). Included in this repertoir of proteins are many of the targets of proposed transmission blocking immunity. It has been reported that An. gambiae can concentrate the bloodmeal by a factor of 1.2 to 1.8 reducing the production of ookinetes when compared to An. freeborni which can not concentrate (Sinden et al. 1996). The mature ookinete is a motile cell that varies from 7 to 18 mm in length and 2 to 4 mm in diameter (Sinden, 1978). Locomotion of the ookinete has been described in different Plasmodium species as a linear or snake-like gliding motion (Freyvogel 1966). Studying P. berghei ookinetes in primary mosquito cell cultures, Speer et al. (1974) described spiral waves on the surface of some ookinetes, especially in the anterior half of the body, which might be involved in ookinete locomotion. Some factors could influence the development, survival and infectivity of the parasite during its residence in the midgut lumen. Eyles (1952) has showed that the parasite development ceases at the ookinete stage unless a macromolecular (non- dialyzable) component is present in the blood meal. Studying the influence of red blood cells on the ability of P. gallinaceum zygotes fertilized in vitro to infect Aedes aegypti Rosenberg et al. (1984) found a linear relationship between erythrocyte density and the number of oocysts up to a 50% hematocrit. Furthermore, they deduced that there are one or more nondialyzable substances (erythrocytic factors) contained in normal erythrocytes, and released by mosquito digestion, that are essential for ookinete invasion of the gut epithelium. When they added trypsin inhibitor to the bloodmeal there was an inhibition of midgut penetration by ookinetes. Recent studies have shown that when mosquitoes are fed with cultured P. falciparum (Ponnudurai et al. 1989) and P. berghei (Sinden 1989) gametocytes, upon dilution with fresh red cells, more oocysts result at initial (low) dilutions whereas further dilution reduces oocyst counts. The involvement of blood factors and/or its digestive products in infectivity has been studied in different parasite-vector models. Using a selected line of An. stephensi, Feldmann and Ponnudurai (1989) found mature P. falciparum ookinetes in the midgut lumen of refractory mosquitoes but no further penetration of the gut epithelium was observed. The reasons for this limited development in non- compatible mosquitoes could be related to digestive function since early initiation of hemoglobin degradation and higher aminopeptidase activity have been described in refractory strains of An. stephensi (Feldmann et al. 1990). It has also been shown that P. gallinaceum develops up to the ookinete stage in the non-compatible mosquito An. stephensi, this development occurring over the same time period and with the same success as in the compatible vector Ae. aegypti. However, P. gallinaceum ookinetes did not escape from the midgut lumen in An. stephensi mosquitoes (Rudin et al. 1991). Possible mechanism inhibiting parasite development involves damage of the parasite by digestive enzymes present in the vector. Trypsin and aminopeptidases are the major proteolytic enzymes involved in blood digestion by female mosquitoes, and are produced by the midgut cells in direct response to blood feeding (Briegel & Lea 1975, Graf & Briegel 1982, Billingsley 1990, Billingsley & Hecker 1991). P. gallinaceum ookinetes 0-10 hr old (i.e. zygote to ookinete transition) were shown to be susceptible to mosquito enzymes in double feeding experiments (Gass 1977) and in vitro damage was observed to cultured ookinetes by proteinases from Ae. aegypti (Gass & Yeates, 1979). However, results found by Shahabuddin et al. (1993) using the same parasite/vector system suggest that the parasite secretes an inactive or partially active chitinase that is activated by a mosquito- produced serine protease. In a recent study Chege et al. (1996) examined the effect of digestive enzymes on the kinetics of P. falciparum ookinete development and oocyst infection rates in An. albimanus, An. freeborni and An. gambiae. Their data indicated that proteolytic enzymes alone do not limit the early stages of sporogonic development in these vector species of Anopheles. Peritrophic layer The peritrophic layer (PL) of the insect midgut forms a cylindrical sheet separating the midgut contents from the single cell-layered midgut endothelium. It is secreted within hours of the blood meal at different rates depending on the mosquito species (Billingsley 1990). Apical secretion granules are present in the midgut cells of Anopheles species, which are released into the periphery of the posterior midgut lumen during the feeding process in response to stretching of the gut wall. The vesicle contents coalesce and then condense to form a compacted PL between 8 and 24 hr (Hecker 1977, Berner et al. 1983). In culicine mosquitoes the PL is formed de novo by the posterior midgut cells in the proventriculus. In A. aegypti, formation starts immediately after the blood meal, but about 12 hr later it is a mature structure (Perrone & Spielman, 1988). The role of the mosquito PL as a barrier to ookinete invasion of the gut wall and pathogenic effects of Plasmodium species upon the vector are controversial. In ultrastructural studies on the interaction of P. falciparum ookinetes with the midgut epithelium of An. stephensi Meis & Ponnudurai (1987) frequentely found parasites trapped in the membrane. They also observed that if the PL was dissected out 36 hr after the blood meal many ookinetes were attached to its external surface. In the same study it was mentioned that the ookinete was capable of penetrating the newly formed, but not the thickened and hardened PL 36 hr after the feeding. A failure to cross this barrier or retarded penetration might increase the length of exposure of ookinetes to mosquito trypsin to which it is known to be sensitive. In contrast, using P. berghei-infected An. atroparvus mosquitoes, Sluiters et al. (1986) concluded that the PL would not function as physical barrier against migrating ookinetes which can pass through fenestrations. Billingsley and Rudin (1992) observed that infectivity of An. stephensi by P. berghei, measured by oocyst counts, was unnaffected by the presence or absence of the PL. However, in A. aegypti infected with P. gallinaceum its presence serves to reduce rather than prevent infection (Ponnudurai et al. 1988, Billingsley & Rudin 1992). These observations suggested to the authors that in compatible vector-parasite combinations, the PL acts as a limiting, rather than absolute barrier to the penetrating ookinete while in incompatible combinations the PL appears to be an absolute barrier to ookinete penetration. To reach the midgut epithelium the ookinetes must first cross the partially or fully formed PL. The PL may act as the recognition site for the penetrating ookinete via lectin- mediated mechanisms. The occurrence of sugar residues which could be involved in vector recognition by the parasite has been demonstrated in the PL of An. stephensi and Ae. aegypti (Berner et al. 1983). Rudin and Hecker (1989) showed the presence of binding sites for different lectins in midguts of P. berghei-infected An. stephensi, with high specificity for N-acetyl-D-galactosamine (GalNAc), and P. gallinaceum-infected Ae. aegypti, with high specificity for N-acetyl-D-glucosamine (GlcNAc). The authors concluded that it seems likely that lectin-binding phenomena play a role in the orientation of the parasites on their way out of the midgut lumen and that the PM and/or glycocalyx may be crucial structures for the penetration of the gut epithelium by the ookinete. This does not exclude the possibility that ookinetes penetrate the PL by an enzymatic process (see below). Ultrastructural observations on Ae. aegypti infected with P. gallinaceum showing an electron dense amorphous material in front of the parasite were consistent with a blockade of parasites within the PL (Sieber et al. 1991). The ookinetes appeared to disrupt the layers of the PL, suggesting an enzymatic mechanism for penetration. These observations were further investigated by Huber et al. (1991) who, using the same vector/parasite system, also suggested a possible role for GlcNAc in the binding of the ookinete to the PL, and demonstrated the presence of chitin in the PL. They observed that mature ookinetes transiently secreted a soluble chitinase, thought to be responsible for the digestion of the PL. Recently, Shahabuddin et al. (1993) reported an inhibition of P. gallinaceum chitinase and a transmission blocking activity of a chitinase inhibitor, allosamidin, on the sporogonic development of P. gallinaceum and P. falciparum in the respective vectors Ae. aegypti and An. freeborni. However, enzymatic mechanisms of penetration may differ in other vector/parasite systems since the PL of An. stephensi appears not to contain chitin (Berner et al. 1983).
Penetration of midgut epithelium Ookinetes found in the epithelial cell layer between 24 and 48 hr post-infection have successfully evaded the obstacles presented by the midgut lumen; however, they still might fail to develop at the normal site of development (Sinden 1984, Ponnudurai et al. 1988). It has been shown that the midgut wall is negatively charged (Houk et al. 1986), but there is no evidence of electrical charge interactions between ookinetes and epithelial cells. It has been a matter for discussion as to whether the ookinete follows an intra- or intercellular route to reach the ultimate site of development and encystment in the outer wall of the midgut epithelium. Intercellular movement of P. gallinaceum ookinetes was first described by Stohler (1957), but Mehlhorn et al. (1980) have found the same parasite in an intracellular position. Recently, Torii et al. (1992a) observed P. gallinaceum ookinetes in both intracellular and intercellular positions in the midgut epithelium of the mosquito Ae. aegypti, which they interpret as that they first enter into the epithelium, then exit into the intercellular space and move to the basal lamina. Garnham et al. (1962) showed that the ookinete of P. cynomolgi bastianelli enters the epithelial cell by liquifying the cell membrane. Davies (1974) again postulated intercellular movement by P. berghei nigeriensis ookinetes. Although describing the ookinete of P. berghei in an intracellular position, as did Garnham et al. (1969), Canning and Sinden (1973) stated that the parasite might also migrate by an intercellular route. More recently, P. yoelii nigeriensis ookinetes have been described to take an intracellular route to the external wall of the midgut (Maier et al. 1987). However, when the same parasite was used to infect An. omorii, an intercellular route was mostly undertaken, although the intracellular occurrence was also observed (Syaffruddin et al. 1991). Meis and Ponnudurai (1987) presented evidence that P. falciparum ookinetes migrate between the epithelial midgut cells. Using a specific monoclonal antibody they also observed a track in the PL, which is related to the shedding of a 25 kD surface protein (Pfs25) during movement. The authors suggested that this protein may bind to receptors on the epithelial cells prior to an intercellular invasion, since it is reported to have epidermal growth factor (EGF)-like domains (Kaslow et al. 1988). It was recently shown that Pfs25 persits in the oocyst wall during parasite development in the mosquito (Lensen et al. 1993). The same group studied the migration of P. falciparum and P. berghei ookinetes through the midgut epithelium in An. stephensi by using ruthenium red staining (Meis et al. 1989). The results of previous studies were confirmed: P. falciparum ookinetes penetrated by intercellular route, but the rodent parasite P. berghei appeared to take an intracellular position, confirming that both mechanisms occur and are species-dependent. In the case of P. berghei a protein of 21 kD (Pbs21) present on the surface of the ookinete (Sinden et al. 1987) could play a role during the intracellular invasion of the midgut epithelium of An. stephensi mosquitoes. It was demonstrated in the midgut of P. berghei infected mosquitoes that expression of Pbs21 was predominantly localized on the ookinete surface one day after the infectious blood meal and thereafter expression declined to a minimum on days 2 and 3, the time of onset of oocyst development (Simonetti et al. 1993). The mode of penetration by ookinetes can perhaps be related to damage of the epithelial lining of the midgut. Intercellular migration may not damage cell membranes, and increased mortality does not occur in P. falciparum-infected An. stephensi and An. gambiae during this period, even with very heavy parasite loads (Meis & Ponnudurai, 1987). Similar observations have been reported in P. gallinaceum-infected Ae. aegypti (Freier & Friedman, 1987). They observed similar mortality rates in infected and uninfected batches of mosquitoes. However, when ookinetes use an intracellular route, as described previously, increased damage to midgut cells might occur, resulting in higher mosquito mortality. This is probably mediated by invasion of the hemolymph by opportunistic gram-negative bacteria and/or microsporidia (Maier et al. 1987, Seitz et al. 1987). Ultimately the ookinete penetrates the basement membrane, but fails to pass through the basal lamina of the midgut adjacent to hemocoele (Sinden 1984). Whether this is due to the inability of the ookinete to penetrate the basal lamina, or to the specific recognition of the lamina and consequent shut- down of the incisive process is not known. Interactions of parasites with vector extracellular matrix proteins (ECM) cannot be discounted (Kaslow et al. 1994b). Oocyst development
Oocyst development is predominantly extracellular (Duncan et al. 1960, Garnham et al. 1969, Howells & Davies 1971, Sinden 1975), but occasionally occurs within the midgut epithelial cell (Vanderberg et al. 1967, Bafort 1971, Beaudoin et al. 1974). The ookinete usually comes to rest beneath the basal lamina 18-24 hr after the infective blood meal (Sinden 1978). It rapidly rounds up between 18 and 72 hr after feeding and the apical complex is resorbed into the oocyst cytoplasm (Garnham et al. 1969). There is some evidence suggesting a significant role of the basal lamina in the development of the ookinete (Kaslow et al. 1994b). It was found that in vitro- cultured ookinetes injected directly into the hemolymph form clusters of oocysts adherent to the basal lamina throughout the hemocoele. Furthermore, binding of ookinetes to artificial surfaces, such as plastic, is enhanced at least 10- fold by addition of various components of basal lamina such as matrigel, collagen IV, and laminin (Warburg & Miller, 1992). The young oocyst is enveloped by a thick plasmalemma that is covered on the hemocoelomic surface by a fibrous basal lamina. Oocysts from the second day onwards are also covered by an amorphous capsule which becomes reduced in thickness at maturity (Vanderberg et al. 1967, Aikawa 1971, Strome & Beaudoin 1974, Sinden 1975). Despite the usual growth of the oocyst under the basal lamina of the midgut, oocyst development is not site specific. Weathersby (1952, 1954, 1960) has shown by injection of gametocytes directly into the hemocoele of susceptible mosquitoes that oocysts would develop to maturity if attached to other parts of the body than the stomach or even if they were floating freely in the hemocoel fluid. In his experiments he used different parasite-host combinations and concluded that the site of oocyst development is probably not a critical factor in the maturation process. Furthermore, the factors that are responsible for the death of parasite in refractory lines are not confined to the stomach wall. These results were supported by those reported by Ball and Chao (1957, 1960, 1961, 1976) who showed that oocysts of P. relictum may develop in vitro away from the intact stomach of the mosquito. The overall results of this series of studies by Ball and Chao demonstrated in vitro development of all stages from ookinetes to fully infective sporozoites without attachment of oocysts to the midgut. However, it was not possible to obtain complete sporogonic development in a single preparation. Rosenberg and Koontz (1984) injected cultured P. gallinaceum zygotes into the hemocoele of Ae. aegypti mosquitoes and observed development of ectopic oocysts in approximately 50% of the mosquitoes, with sporozoites being found in the salivary glands. These observations suggest that oocyst metabolism is not dependent upon direct transfer of nutrients from the midgut epithelium. Occasional intracellular oocyst development has been reported for P. berghei in An. stephensi and An. quadrimaculatus (Vanderberg et al. 1967). The same localization was described in P. vinckei by Bafort (1971) who concluded that both mechanical pressure and physiological mechanisms play a role in the movement of oocysts to the hemocoelomic surface. Studying the sporogonic development of P. berghei in An. stephensi, Beaudoin et al. (1974) found oocysts developing ectopically within the midgut epithelium following normal infection, eventually emptying their sporozoite content into the tissue itself or the midgut lumen. In addition, they observed no morphological and structural abnormalities in the luminal parasites which displayed good viability. In contrast to P. berghei, no ectopic development was seen in P. falciparum-infected An. stephensi mosquitoes (Meis et al. 1992b), confirming previous results observed with P. falciparum in naturally infected An. gambiae (Sinden & Strong 1978). Recent reports described an enhancement of oocyst development in vitro for P. berghei (Syaffruddin et al. 1992), P. gallinaceum (Warburg & Miller, 1992) and P. falciparum (Warburg & Schneider, 1993), when insect cell lines were added into the culture medium. From these observations it appears that nutritional or other regulatory requirements of the developing parasite can be met without a direct contact with midgut epithelium or haemolymph. The question of how the oocyst is supplied with nutritive material is an intriguing one. Little information is available on the uptake and source of nutrients for oocyst development. It is assumed that in vivo the source of nutrients is the hemolymph. Mack and Vanderberg (1978) analyzed hemolymph of An. stephensi collected from uninfected and P. berghei-infected mosquitoes at different stages of the parasite development. It was found that four days after the blood meal the osmotic pressure and the specific gravity were lower in infected mosquitoes compared with uninfected ones. The difference, however, was attributed to indirect effects of the quality of the ingested blood meal. These studies were complemented by analysis of the concentration of free amino acids in the hemolymph collected in similar conditions with results showing significantly lower concentrations in infected mosquitoes with decreases in valine and histidine, and a total loss of detectable methionine suggesting it is incorporated (Mack et al. 1979). This difference could be due to the utilization of some of these amino acids by the developing oocyst as suggested by Ball and Chao (1976) who analyzed the uptake of amino acids by P. relictum oocysts in vitro, comparing growth of uninfected and infected guts of Culex tarsalis in Grace's insect culture medium. They found significant decreases in the concentration of certain amino acids including arginine, asparagine, proline and histidine, and less marked decrease in concentrations of others like methionine, valine, leucine and isoleucine. From these studies it appears that the reduced amount of free aminoacids in the hemolymph is due to oocyst metabolism. Autoradiographic studies with P. gallinaceum in Ae. aegypti mosquitoes indicated that ^3H-leucine is uniformly incorporated throught the oocyst within 15 min of injection into hemocoele (Vanderberg et al. 1967).
Sporogony With increasing maturation the oocyst undergoes considerable cytoplasmic subdivision. Initially the plasmalemma forms invaginations and clefts that penetrate even deeper into the cytoplasm, thus subdividing the cell (Vanderberg et al. 1967, Terzakis 1971, Posthuma et al. 1988). In a transmission electron microscopy study of P. falciparum oocysts it was suggested that cleft formation was due to dilation of endoplasmic reticulum (Sinden & Strong 1978). Using immunogold labelling technique during sporogonic stage of the same parasite, Posthuma et al. (1988) considered the latter explanation unlikely. With increasing activity the cytoplasmic clefts become extended and the expanding vacuolar space more pronounced leading to the sporoblast formation. Along the clefts sporozoites are formed by a budding process at the surface of the limiting membrane (Vanderberg et al. 1967, Howells & Davies 1971, Canning & Sinden 1973, Sinden & Strong 1978). As the sporozoite continues to bud off, a nucleus and various cytoplasmic components are passed into it from the sporoblast. The membranes of the developing sporozoite pellicle are formed and other organelles like microtubules and rhoptries become discernible (Vanderberg et al. 1967, Sinden & Garnham 1973). When sporogony is completed (about 10-12 days after the infective feed), the oocyst is filled with sporozoites and one or more residual bodies (Sinden 1984). Estimations of number of sporozoites per oocyst have ranged widely. Garnham (1966) reported that the number in a single P. vivax oocyst varies from 1,000 to 10,000. Pringle (1965) estimated that a single oocyst of P. falciparum contains nearly 10,000 sporozoites. Studies carried out with mosquitoes fed on infected volunteers from Thailand showed a mean count of approximately 3,700 sporozoites per oocyst for P. vivax and 3,400 for P. falciparum, whereas for P. cynomolgi-infected mosquitoes a single oocyst contained about 7,500 (Rosenberg & Rungsiwongse 1991). Dependent upon species, the mature sporozoite varies from 9 to 16.5 mm in length and from 0.4 to 2.7 mm in diameter; aberrant forms have been described up to 40 mm long (Sinden 1978). So far, studies on synthesis and expression of proteins during sporogonic development have foccused on a polypeptide called the circumsporozoite protein (CSP) found on the mature sporozoite. Observations on the origin of CSP and its distribution through the mosquito stage were reported by several authors. It is now well established that these proteins are synthesized in maturing oocysts of different Plasmodium species from 6-7 days after the infective blood meal, before sporozoites are visible (Nagasawa et al. 1987, 1988, Posthuma et al. 1988, Hamilton et al. 1988, Boulanger et al. 1988, Torii et al. 1992b, Meis et al. 1992a). At this stage, CSP is present on the plasmalemma and at various sites within the cytoplasm and endoplasmic reticulum of the sporoblast. When the sporozoites bud from the sporoblast they are already covered with CSP which is also found in salivary gland sporozoites (Yoshida et al. 1981, Santoro et al. 1983, Tsuji et al. 1992). Humoral encapsulation of oocysts, which in malaria infected mosquitoes is known as Ross' black spores, has been described in P. berghei nigeriensis and P. vivax and seems to occur mainly in older oocysts which have begun to produce sporozoites (Sinden & Garnham, 1973). This phenomenon was studied in a selected line of An. gambiae that encapsulates different Plasmodium species (Collins et al. 1986). The authors demonstrated that refractoriness is manifested by melanization of the ookinete after its passage through the midgut epithelium. Paskewitz et al. (1989) localized phenoloxidase activity in the basal lamina of the epithelial cells of both encapsulating and susceptible mosquitoes prior to blood feeding. However, after an infective blood meal, this activity was still observed close to invading ookinetes in refractory mosquitos but it was reduced or absent in susceptible mosquitoes. When the non-compatible vector An. gambiae was fed with ookinetes of P. gallinaceum, invasion of the midgut epithelium by the ookinetes occurred but oocysts were infrequentely formed. Using the same system Vernick et al. (1989) and Vernick and Collins (1989) tried to elucidate mechanisms involved in vector-parasite incompatibility by injecting in vitro- cultured ookinetes into the hemocoel of mosquitoes and monitoring parasite development using specific rRNA probes. As no differences were found between susceptible and refractory mosquitoes the authors suggested that the specific lytic factor(s) in the refractory line are intracellular. The sporozoite and salivary gland invasion
The motile sporozoites emerge into the hemocoele through holes from an area of weakness in the oocyst wall. Holes are possibly produced by a combination of the muscular action of the gut wall and the activity of the sporozoites (Sinden 1978, Meis et al. 1992b). Within the hemocoel the sporozoites are distributed throughout the mosquito and can initially be found in many parts of the insect, even in the maxillary palps; within a day or two of their release from oocysts the sporozoites invade the salivary glands where they accumulate and remain until delivery (Vaughan et al. 1992). Thus, sporozoites do not adhere to most tissues, except for the salivary glands and rarely the midgut wall, hemocytes or thoracic muscles (Vanderberg 1974, Sinden 1975, 1978, Golenda et al. 1990, Vaughan et al. 1992). The latter observation, especially in heavily infected mosquitoes, could be associated with an impairment of flight activity in malaria-infected An. stephensi vectors demonstrated by some authors (Rowland & Boersma 1988). It has been estimated that in mosquitoes fed on individuals with naturally acquired P. vivax about 850 sporozoites per oocyst reached the glands (Sattabongkot et al. 1991) which follows, by calculation, that approximately 23 % of all P. vivax sporozoites released into the hemocoel subsequentely reach the salivary glands (Rosenberg & Rungsiwongse 1991). Vaughan et al. (1992), using regression analysis, calculated the approximately 650 salivary gland sporozoites were produced per oocyst and reported that virtually all oocyst infections produced salivary gland infections in An. gambiae infected with P. falciparum by membrane feeding. The same group has found a similar number by studying the sporogonic development of cultured P. falciparum in six species of laboratory-reared Anopheles mosquitoes (Vaughan et al. 1994). This is in contrast to observations on wild-caught An. gambiae from Burkino-Faso (Lombardi et al. 1987) and western Kenya (Beier et al. 1990) where sporozoites failed to enter the salivary glands in 43% and 10% of infected mosquitoes, respectively. However, when sporozoites from P. gallinaceum oocysts were injected into Ae. aegypti female mosquitoes only 6.5 to 10.4% of inoculated sporozoites invaded the salivary glands. Interestingly, injected salivary gland sporozoites did not reinvade the glands (Touray et al. 1992). Recently, a laboratory study on An. tesselatus mosquitoes infected with different isolates of P. vivax and P. falciparum from patients living in Sri Lanka showed that approximately 15% of mosquito batches in which oocysts developed failed to produce salivary gland sporozoites (Gamage-Mendis et al. 1993). This discrepancy between naturally and laboratory-infected mosquitoes could be attributed to different environmental conditions or mixed mosquito populations. It could be expected sporozoites would elicit a humoral response in the mosquito by activation of the prophenoloxidase cascade and as a result be killed by melanization. However, sporozoites in the hemocoele are rarely seen to be melanized (Brey 1991). Sporozoites might be protected from mosquito defense reactions against non-self' by antigen sharing. This has been demonstrated as a potential mechanism for avoidance of mosquito defence reactions by microfilariae of Brugia pahangi (Maier et al. 1987). Immunoreactivity to CSP was observed on the midgut wall of mosquitoes infected with P. falciparum (Boulanger et al. 1988) and P. yoelii (Beaudoin et al. 1990). The latter authors also detected reactivity on uninfected midguts and suggest the presence of a common determinant between the parasite and the mosquito. The migration of sporozoites from oocysts to salivary glands could be active (in contact with basal lamina), passive (in suspension) or both. If active, the parasite would use a gliding motility to move across the basal lamina and reach the gland. Vanderberg (1974) described circular gliding and attached waving locomotion and movement in sporozoites of different species when bovine serum albumin was added to Medium 199. Sporozoites that move over a substratum in vitro leave behind trails of CS proteins (Stewart & Vanderberg 1988, 1991, 1992). Soluble CSP was shown to be present throughout the hemocoel (Robert et al. 1988). Beier et al. (1992a) concluded that the release of CSP by sporozoites is a normal but complex mechanism that they interpreted to be associated with sporozoite survival in the host, is not site- specific and it would be regulated in response to background levels of soluble CSP in the environment (negative feedback mechanism). This idea is supported by previous results showing that the parasites cease to synthesize CSP during their journey through the hemolymph but shedding still occurs (Posthuma et al. 1988, 1989). There is no evidence yet for chemotaxis but the congregation of sporozoites in the vicinity of the glands before invasion may support, according to some workers, a tactile mechanism (Golenda et al. 1990, Meis et al. 1992b). Several polysacharides have been identified that may orient protozoa towards or away from a stimulus (Van Houten 1988). The selective invasion of mosquito salivary glands by malarial parasites is not well understood. Specific recognition of the glands was shown in P. knowlesi. Oocysts developed normally on the gut of An. freeborni but sporozoites were never found in salivary glands. When glands from the susceptible vector An. dirus were implanted into the abdomen, they did become infected (Rosenberg 1985). These experiments demonstrated that the salivary glands themselves determined specificity. Although invasion of salivary glands seems to require specific recognition, the mechanism by which sporozoites recognize, attach to, and penetrate the glands remains to be determined. Sporozoites preferentially invade the medial lobe and the distal portions of the lateral lobes of the salivary glands where the salivary duct is not chitinous in Anopheles species (Sterling et al. 1973, James & Rossignol 1991, Ponnudurai et al. 1991). Hence, the occurrence of a specific receptor-mediated invasion by sporozoites is plausible. Perrone et al. (1986) used lectins to characterize carbohydrate moieties on the basal lamina of Ae. aegypti salivary glands. As the median and distal lateral lobes bound a common lectin, RCA 120, whose substrate is b-D- galactose, the authors suggested that sugars that bind this lectin serve as candidate residues to which sporozoites may attach. In contrast, the sporozoite coat binds some lectins with very low efficiency (Schulman et al. 1980, Turner & Gregson 1982). However, in a recent scanning electron microscopic study Meis et al. (1992b) localized P. falciparum sporozoites in proximal and distal parts but were unable to identify any specific regions on the glands where sporozoites penetrate. These authors also showed sites where the sporozoites have pierced the basal lamina, which probably explains the presence of CSP on the basal lamina induced by shedding during penetration (Posthuma et al. 1989) and the presence of immunoreactive spots of 1-2 mm on the surface of infected glands (Golenda et al. 1990). Recently, Touray et al. (1994) developed an in vivo salivary gland invasion assay and have found that anti-salivary gland antibodies, sulfated glycosaminoglycans and some lectins, particularly Suc-WGA, block invasion of sporozoites. Although the mechanism of blocking is not yet known, those lectins that blocked invasion bound to salivary glands but did not bind to sporozoites. Beier et al. (1992a) and Beier (1993) proposed that as sporozoites invade the salivary glands, the build-up of CSP is the signal for sporozoites to halt their active motility, and thus their release of CSP (down-regulation). The involvement of CSP and other proteins distinct from CSP such as PySSP2 (Charoenvit et al. 1987) and PfSSP2 (Robson et al. 1988, Rogers et al. 1992a,b) in binding of sporozoites to the basal lamina of mosquito salivary glands is speculative. As they share similarities in their structure they might be related to this process since it has been suggested region II of CSP is involved in hepatocyte invasion (Cerami et al. 1992a,b, Pancake et al. 1992). Rosenberg (1985) and King (1988) have suggested that sporozoites invade the salivary gland cells using a mechanism similar to invasion of erythrocytes by merozoites although there is no evidence of parasitophorous vacuole membrane formation observed in other stages of the parasite cycle (Sinden & Strong 1978, Meis & Verhave 1988). Membrane-limited vacuoles beneath the plasma membrane in An. stephensi distal salivary gland cells invaded by P. berghei have been described (Sterling et al. 1973). Penetration could also involve an unusual intercellular route as suggested by Golenda et al. (1990) who detected CSP in the region between cells of the median lobe of the gland. Posthuma et al. (1989) observed many sporozoites on the basal side of the cells, but also found trail-like CSP immunoreactivity at the lateral space between the cells. After penetration, sporozoites are present in bundles in the accini of gland cells in both proximal and distal areas. Most probably, the sporozoites which are present in proximal areas of the glands are unable to reach the draining duct because of the chitinized layer in that area whereas distally localized sporozoites reach the draining duct via the large unchitinized collecting tube (Meis et al. 1992b). This explanation would not be valid for Aedes species since the salivary ducts are lined with chitin and extend the full length of the glands (James & Rossignol 1991). Penetration by sporozoites could cause pathological vesiculation and cytoskeletal changes in salivary gland tissues as the infected cells are often deformed and swollen (Sterling et al. 1973, Maier et al. 1987). The efficiency of salivary gland invasion is poorly understood. It has been estimated that the median sporozoite load in the glands is less than 10,000 in colonized or wild Anopheles species (Shute 1945, Pringle 1966, Wirtz et al. 1987, Beier et al. 1991b, Sattabongkot et al. 1991) or slightly higher (Ponnudurai et al. 1991). The development of infectivity by the sporozoites appears to be asynchronous, in some cases taking place in the hemocoele, while in others not occuring until after they have invaded the salivary glands. Thus, it seems to be time-dependent rather than site-dependent (Vanderberg 1975, Daher & Krettli 1980). It was demonstrated in P. berghei-infected mice that populations of salivary gland sporozoites were more than 10,000 times as infective to the vertebrate host as populations of oocyst sporozoites from the same mosquitoes (Vanderberg 1975). Touray et al. (1992) found that as few as 10-50 salivary gland P. gallinaceum sporozoites were required to induce infection in chickens compared to 5,000 oocyst sporozoites. Sporozoite infectivity increases with time during the first week after the invasion of the salivary gland (Vanderberg 1975). Degeneration of sporozoites is not frequent in nature as observed by Barber (1936) who studied anophelines collected in Mediterranean areas. When degeneration occurred in a salivary gland or a lobe of a gland, in another gland or lobe the sporozoites were normal or in a different stage of degeneration. The number of sporozoites injected into the tissue or capillary of the vertebrate is very small compared to that found in the salivary glands. It has been estimated, by employing different methods, that each bite delivers fewer than 50 sporozoites with a tendency for most sporozoites to be ejected in the first droplets of saliva (Vanderberg 1977, Rosenberg et al. 1990, Ponnudurai et al. 1991, Beier et al. 1991a,b, Beier et al. 1992b, Li et al. 1992). Although a correlation between salivary gland sporozoite load and sporozoite inoculum has been reported (Rosenberg et al. 1990) ejection of sporozoites is probably a random process, more related to the architecture of the salivary gland duct system than to the number of sporozoites in this organ (Ponnudurai et al. 1991). Some studies reported that P. falciparum-infected mosquitoes deliver sporozoites in an unpredictable fashion, sometimes not at all (Ponnudurai et al. 1991), and others transmit inconsistently (Rickman et al. 1990). Clumping of sporozoites has been reported in infected salivary glands (Sterling et al. 1973) and clusters of sporozoites were detected after delivery when An. stephensi mosquitoes infected with P. falciparum were allowed to feed through fresh mouse skin (Ponnudurai et al. 1991). It has been observed that salivary glands are not depleted of sporozoites even in vectors that feed up to 15 times (Shute 1945), which allows infected mosquitoes to remain potentially infectious for life. The low sporozoite inoculum and the low entomological inoculation rates in natural conditions (Mendis et al. 1990a, Gordon et al. 1991) demonstrates the high efficiency with which injected sporozoites will develop malaria.
Factors affecting parasite development to the mosquito The process of infection of mosquitoes is exceedingly complex and regulated by a range of factors originating from the parasite, the vertebrate host and the mosquito vector, and from the interactions between all three (Sinden 1991). Many of these and other factors are known to affect fertilization and subsequent stages of parasite development, thus having great influence on transmission of the disease. To have an idea, when sporogonic development of cultured P. falciparum was evaluated in six species of Anopheles mosquitoes there was a total loss of approximately 31,600-fold in the parasite population from macrogametocyte to oocyst stage (Vaughan et al. 1994).
a) Host location and feeding behaviour - Of primary importance in malaria transmission is the proportion of mosquito feeds taken from humans and the proportion of these feeds taken from infected individuals. In nature a vector needs to survive longer than the sporogonic period after taking an infective blood meal; during this period the mosquito probably takes blood meals every 2-3 days, depending on its gonotrophic cycle and the availability of breeding sites (Ponnudurai et al. 1989). Parasitemic hosts tend to be sick and often less irritated by mosquito feeding. Additionally, the thrombocytopenia which is commonly associated with blood-borne parasitic diseases leads to facilitation of vessel location, resulting in increased feeding success by mosquitoes on parasitemic hosts (Rossignol et al. 1985). Salivary glands and saliva contain a whole range of components with pharmacological activities important for blood feeding success and subsequent bloodmeal processing, including anticoagulants, anti-inflammatory, vasodilatory and immunosuppressive compounds (Ribeiro et al. 1984, 1989, Ribeiro 1987, Titus & Ribeiro 1990, James & Rossignol 1991). Rossignol et al. (1984) concluded that the lesions caused by P. gallinaceum sporozoites in the salivary glands of Ae. aegypti result in reduced levels of salivary apyrase. Mosquitoes deficient in salivary apyrase experience difficulty in locating host blood and engorging; they therefore probe more often and may attempt to feed on several hosts in succession (Rossignol et al. 1984, 1986, Ribeiro et al. 1985). Li et al. (1992) demonstrated that probing time of P. berghei-infected An. stephensi is not affected by sporozoite invasion of salivary glands. Blood meal size may influence infectivity since it determines the number of gametocytes ingested and therefore subsequent infection. It has been shown that larger An. dirus females took larger bloodmeals by artificial feeding with cultured P. falciparum and developed significantly more oocysts (Kitthawee et al. 1990). In a field study, Lyimo and Koella (1992) reported that the proportion of An. gambiae mosquitoes infected with P. falciparum during a blood meal was independent of size but the number of oocysts harboured by infected mosquitoes increased with size of the mosquito.
b) Gametocyte carriers/transmission blocking immunity - Malarial infections induce host responses to both asexual and sexual stage malaria parasites that may modulate gametocyte infectivity. Great heterogeneity in infectiousness of different carriers has been noted, with apparently poor correlation between infectiousness and gametocyte density (Graves et al.1988). However, it remains unclear whether symptomatic and asymptomatic asexual infections differ in their ability to influence gametocyte infectivity. It has been reported that asymptomatic P. falciparum patients were more infectious than symptomatic patients (Boyd & Kitchen 1937, Jeffery & Eyles 1955, Muirhead-Thomson 1957, Carter & Graves 1988) while, in contrast, another report suggested that asymptomatic and symptomatic P. malariae patients were equally infective (Young & Burgess 1961). Specific and non-specific responses are believed to modulate parasite transmission. The induction of specific immunological responses to Plasmodium shows marked heterogeneity within human populations (Mendis et al. 1987, Graves et al. 1988, Targett 1990, Snow et al. 1993). The sexual stages can induce trasmission blocking immunity (TBI) and effective targets include antigens identified on the surface of the macro- and/or microgemetes ( pre-fertilization'antigens) as well as antigens present on the surface of the gamete, zygote and ookinete ( post-fertilization' antigens). Antibodies to pre-fertilization' antigens of P. falciparum such as Pfs230, Pfs48/50 and Pfs16 have been detected in humans (Graves et al. 1988, Premawansa et al. 1994, Hogh et al. 1994). A significant association between lacking of infectivity of P. falciparum gametocyte carriers and recognition of epitope IIa on Pfs48/50 by antibodies in their sera has been observed (Graves et al. 1992). Naturally acquired TBI to P. vivax sexual stage antigens has also been demonstrated (Mendis et al. 1987, 1990b, Mendis & Carter 1991, Ranawaka et al. 1988, Goonewardene et al. 1990, Gamage- Mendis et al. 1992) and appears to play a role on transmission of the disease (de Zoysa et al. 1988). In P. vivax malaria antibodies, at low concentrations, can also have a transmission-enhancing effect on infectivity of malarial parasite to mosquitoes (Mendis et al. 1987, Peiris et al. 1988, Naotunne et al. 1990, Gamage-Mendis et al. 1992). A recent report described TBI in P. vivax malaria when antibodies raised against a peptide blocked parasite development in the mosquito An. tesselatus (Snewin et al. 1995). As shown in a variety of Plasmodium species, within the mosquito vector antibody to the pre-fertilization' antigens may prevent fertilization by any of four mechanisms: (1) the agglutination of macro/microgametes limiting their mobility; (2) antibody coating of macro/microgamentes inhibiting cell-cell recognition; (3) opsonization in the bloodmeal or (4) complement dependent/independent lysis (Gwadz 1976, Carter & Chen 1976, Carter et al. 1979, 1985, 1990, Kaushal et al. 1983a,b, Rener et al. 1983, Harte et al. 1985a, Vermeulen et al. 1985b, Grotendorst et al. 1986, Grotendorst & Carter 1987, Quakyi et al. 1987, Peiris et al. 1988, Premawansa et al. 1990). Cellular responses are also involved in TBI and may have some influence on parasite infectivity (Harte et al. 1985b, Mendis et al. 1990b, Riley & Greenwood 1990, Goonewardene et al. 1990). Immunity to `post-fertilization' antigens such as Pfs25, Pbs21, Pgs25 and Prs25 also plays an important role on transmission of the disease by suppressing parasite infectivity at different stages of its development in the mosquito as demonstrated by many workers in different Plasmodium species (Grotendorst et al. 1984, Vermeulen et al. 1985a, Sinden et al. 1987, Winger et al. 1988, Fries et al. 1989, Carter & Kaushal 1984, Carter et al. 1989a,b, Kaslow et al. 1991, 1992, 1994b, Foo et al. 1991, Sieber et al. 1991, Tirawanchai et al. 1991, Duffy et al. 1993, Paton et al. 1993, Ranawaka et al. 1993, 1994). The mechanisms of blockade could be the same four described above and/or the antibody may act by damaging the parasite surface coat (Ponnudurai et al. 1987). Non-specific responses to the asexual stages are believed to modulate parasite trasmission (Naotunne et al. 1991, Kwiatkowski 1992). Numerous non-specific factors may correlate with changes in gametocyte infectivity. Acute phase reactants like C-reactive protein (CRP), which are non-specific indicators of inflammatory activity, are elevated in patients with P. falciparum malaria (Ree 1971, Naik & Voller 1984, Chagnon et al. 1992). Some cytokines such as interferon (IFN-g), tumor necrosis factor (TNF-a) and interleukin 6 (IL- 6) are elevated in sera fom patients with P. falciparum and P. vivax malaria (Grau et al. 1989, Kern et al. 1989, Kwiatkowski et al. 1990, Mendis et al. 1990c, Karunaweera et al. 1992). Both IFN-g and TNF-a appear to cause a transient but marked drop in the infectivity of gametocytes to mosquitoes due to the intraerythrocytic killing of parasites (Naotunne et al. 1991, Karunaweera et al. 1992). However, a recent study has shown no elevation in blood levels of cytokines IL-2, IL-6, TNF-a and IFN-g nor reactive nitrogen intermediates (Hogh et al. 1994). The authors suggested that this could be explained by the inability of asymptomatic gametocyte carriers, unlikely to harbour high asexual parasitaemias, to promote the responses. The concept that anti-sexual stage immunity may regulate infection of the moquito vector by gametocyte-infected malarial blood has gained considerable support and must be considered for the development of malaria trasmission blocking vaccines (Kaslow et al. 1992).
c) Antibodies against sporozoites - There is evidence that naturally acquired or experimentally elicited anti-sporozoite antibodies ingested by mosquitoes may affect the dynamics of the sporogonic development in the vector. Several studies with P. falciparum- infected Anopheles species (Vaughan et al. 1988, Beier et al. 1989, Hollingdale & Rosario 1989) showed that (1) ingested human CSP antibodies were detected in the blood meal of field collected mosquitoes up to 36 hr after feeding, (2) antibodies crossing the midgut into hemocoel persist from 4 to 36 hr post-infection in hemolymph, (3) ingested CSP antibodies on day 5 after infection bound to developing oocysts, (4) enhancement of the sporozoite production, (5) ingestion of CSP antibodies on day 10 after feeding had no effect on oocyst maturation or sporozoite production, (6) contact between CSP antibodies and sporozoites in the hemocoel did not block sporozoite invasion of salivary glands, (7) exposure to CSP antibody increased sporozoite infectivity and (8) human IgG antibodies were present on salivary gland sporozoites from field-collected mosquitoes. It has been recently demonstrated that antibodies to P. gallinaceum CSP prevent sporozoites from invading salivary glands of Ae. aegypti (Warburg et al. 1992). Ponnudurai et al. (1989) did not find any influence of anti-sporozoite antibodies on the number of salivary gland sporozoites but concluded that a second blood meal, with or without antibody, simply functions as a nutritional stimulus for faster oocyst maturation. However, when transmission blocking antibodies anti-Pbs21 (a surface antigen present on the surface of P. berghei zygote/ookinete) were added to second bloodfeeds at different stages of parasite development in the mosquito, a significant reduction in oocyst intensity but no detectable change in prevalence occurred. Furthermore, at all times anti-Pbs21 reduced sporozoite number in the thorax but highest gland intensities were obtained when the second bloodfeed was given on day 4 (Ranawaka et al. 1993). These results were interpreted as two opposing roles of second bloodfeeds containing trasmission blocking antibody: (1) inhibition of parasite development and (2) the supply of nutrients which permit more sporozoites to be produced by each oocyst. Despite some controversy these results potentially have significant implications for natural malaria transmission and for a possible vaccine development.
d) Anti-mosquito antibodies - In addition to anti-parasite antibodies, it has been tested experimentally the effect on malaria trasmission of antibodies raised against parts of the mosquito which could be included in a malaria vaccine. Ramasamy and Ramasamy (1990), studying the P. berghei/An. farauti model, found that mosquitoes feeding on mice immunised with midgut antigens exhibited a reduction in mosquito infection rates. Similar results were reported by Billingsley et al. (1990) using monoclonal antibodies produced against mosquito midgut tissue in P. berghei/An. stephensi system.
e) Genetic manipulation of the vector - Experiments with refractory lines of An. gambiae (Collins et al. 1986) and studies on the effects of broad antimicrobial and antiparasitic components, e.g. magainins and cecropins (Gwadz et al. 1989), showed that it may be feasible to induce effective disruption in the normal development of Plasmodium species in the vector by the introduction and expression of appropriate genes into the mosquito genome. Two types of useful target genes can be used in transgenic mosquitoes. First, those that render populations vulnerable to subsequent control measures, such as insecticide susceptibility or temperature sensitivity, and second, those that interrupt disease transmission by replacing vector with non-vector forms (Crampton et al. 1990, Kidwell & Ribeiro 1992, Crampton 1994). Although many technical, and perhaps ethical, problems associated with the wild-release of transgenic insects have yet to be overcome, the potential of this technology has received greater attention recently (Brey 1991, Coluzzi 1992, Collins 1994, Curtis 1994). Introduction and expression of genes coding for antibodies against target antigens present on the ookinete surface into the mosquito embryos is one of the possibilities to examine the potential of this technology (Crampton et al. 1993).
f) Anti-malarial drugs - Sub-therapeutic doses of antimalarial drugs have been reported to enhance infectivity of Plasmodium species to their vectors (Shute & Maryon 1954). Additionally, numerous compounds including chloroquine (Wilkinson et al. 1976), sulphamethoxazole-trimethroprim (Wilkinson et al. 1973), pyrimethamine (Shute & Maryon 1951), Fansidar (Carter & Graves 1988) and Berenil (Ono et al. 1993) have been suggested to induce gametocyte formation but no influence of chloroquine (Jeffery et al. 1956, Smalley 1977, Chutmongkonkul et al. 1992, Hogh et al. 1994) and Fansidar (Hogh et al. 1994) on gametocyte infectivity was observed by some investigators. It was demonstrated that pyrimethamine- and halofantrine-treated gametocytes of P. falciparum are more infective to An. stephensi mosquitoes than untreated controls (Chutmongkonkul et al. 1992). Other studies examined the effects of some schizontocidal agents on the sporogonic cycle of P. falciparum and P. berghei in anopheline mosquitoes (Coleman et al. 1988, do Rosario et al. 1988). It was found that chloroquine, when fed during late sporogony (10-12 days post-infection), may increase the vectorial capacity of some mosquito species. The effects of chloroquine on the infectivity of chloroquine-sensitive and -resistant populations of P. yoelii nigeriensis to An. stephensi mosquitoes were studied by Ichimori et al. (1990). The results showed an enhancement of infectivity in sensitive strains but no effect was detected in resistant clones and sublines. Chloroquine use and the subsequent development of resistance over the past years is associated with an increasing human malaria infectiousness (Lines et al. 1991) which may be indirect effects of parasitaemia on the host. The sporontocidal activity of chloroquine, halofantrine and pyrimethamine was evaluated by administration to An. stephensi mosquitoes, either in the first bloodmeal containing P. falciparum gametocytes from in vitro cultures, or in the second, parasite-free bloodmeal, given four days after infection. A sporontocidal effect was observed only when pyrimethamine was administred with the infective bloodmeal (Chutmongkonkul et al. 1992). It has been demonstrated recently an inhibitory action of the anti- malarial Atovaquone (566C80) against ookinete, oocysts and sporozoites of Plasmodium berghei in An. stephensi (Fowler et al.1994, 1995). Acknowledgments Amauri Braga Simonetti was supported by a scholarship from CNPq (Brazil) during his PhD in the Infection and Immunity Section, Department of Biology at Imperial College of Science, Technology and Medicine (London, U.K.). I would like to acknowlegde Professor Robert E Sinden for his guidance and helpfull advices which made possible this review. References Aikawa M 1971. Plasmodium: The fine structure of malarial parasites. Exper Parasitol 30: 284- 320. Aikawa M Atkinson CT, Beaudoin LM, Sedegah M, Charoenvit Y, Beaudoin R 1990. Localization of CS and Non-CS Antigens in the Sporogonic Stages of Plasmodium yoelii. Bull WHO 68: 165-171. Alano P 1991. Plasmodium sexual stage antigens. Parasitol Today 7: 199-203. Alano P, Carter R 1990. Sexual differentiation in malaria parasites. Ann Rev Microbiol 44: 429-449. Bafort JM 1971. The biology of rodent malaria with particular reference to Plasmodium vinkei vinkei Rhodain 1952. Ann Soc belge de Med Trop 51: 1-204. Ball G H, Chao J 1957. Development in vitro of isolated oocysts of Plasmodium relictum. J Parasitol 43: 409-412.
Ball GH, Chao J 1960. In vitro development of the mosquito phase of Plasmodium relictum. Exp Parasitol 9: 47-55. Ball GH, Chao J 1961. Infectivity to canaries of sporozoites of Plasmodium relictum developing in vitro. J Parasitol 47: 787-790. Ball GH, Chao J 1976. Use of amino acids by Plasmodium relictum oocysts in vitro. Exp Parasitol 39: 115-118. Barber MA 1936. Degeneration of the sporozoites of the malaria parasite in anopheline mosquitoes in nature and its relation to the transmission of malaria. Am J Hyg 24: 45-56. Beaudoin RL, Strome CPA, Tubergen TA 1974. Plasmodium berghei berghei: ectopic development of the ANKA strain in Anopheles stephensi. Exp Parasitol 36: 189-201. Beier JC 1993. Malaria sporozoites: survival, transmission and disease control. Parasitol Today 9: 210-215. Beier JC, Davis JR, Vaughan JA, Noden BH, Beier MS 1991a. Quantitation of Plasmodium falciparum sporozoites transmitted in vitro by experimentally infected Anopheles gambiae and Anopheles stephensi. Am J Trop Med Hyg 44: 564-570. Beier JC, Onyango FK, Ramadhan M, Koros JK, Asiago C.M, Wirtz RA, Koech DK, Roberts CR 1991b. Quantitation of malaria sporozoites in the salivary glands of wild afrotropical Anopheles. Med Vet Entomol 5: 63-70. Beier JC, Oster CN, Koros JK, Onyango FK, Githeko AK, Rowton E, Koech DK, Roberts CR 1989. Effect of human circumsporozoite antibodies in Plasmodium -infected Anopheles (Diptera: Culicidae). J Med Entomol 26: 547- 553. Beier JC, Perkins PV, Koros JK, Onyango FK, Gargan TP, Wirtz RA, Koech DK, Roberts, CR 1990. Malaria sporozoite detection by dissection and ELISA to assess infectivity of afrotropical Anopheles (Diptera: Culicidae). J Med Entomol 27: 377-384. Beier JC, Vaughan JA, Madani A, Noden, BH 1992a. Plasmodium falciparum: release of circumsporozoite protein by sporozoites in the mosquito vector. Exp Parasitol 75: 248-256. Beier MS, Davis JR, Pumpuni CB, Noden BH, Beier JC 1992b. Ingestion of Plasmodium falciparum sporozoites during transmission by anopheline mosquitoes. Am J Trop Med Hyg 47: 195-200. Berner R, Rudin W, Hecker H 1983. Peritrophic membranes and protease activity in the midgut of the malaria mosquito, Anopheles stephensi (Liston) (Insecta: Diptera) under normal and experimental conditions. J Ultrastruct Res 83: 195-204. Billingsley PF 1990. The midgut ultrastructure of haematophagous insects. Ann Rev Entomol 35: 219-248.
Billingsley PF, Hecker H 1991. Blood digestion in the mosquito, Anopheles stephensi Liston (Diptera: Culicidae): activity and distribution of trpsin, aminopeptidase, and alpha-glucosidase in the midgut. J Med Entomol 28: 865-871. Billingsley PF, Rudin W 1992. The role of the mosquito peritrophic membrane in bloodmeal digestion and infectivity of Plasmodium species. J Parasitol 78: 430-440. Billingsley PF, Winger L, Simonetti AB, Sinden REE 1990. Monoclonal antibdies produced against mosquito midgut tissue. VII International Congress of Parasitology (COPA), Paris. Boulanger N, Matile H, Betschart B 1988. Formation of the circumsporozoite protein of Plasmodium falciparum in Anopheles stephensi. Acta Trop 45: 55- 65. Boyd MF, Kitchen SF 1937. On the infectiousness of patients infected with Plasmodium vivax and Plasmodium falciparum. Am J Trop Med Hyg 17: 253- 262. Brey PT 1991. The Taming of the Anopheles - Current trends in malaria vector research. Res Immunol 142: 712-722. Briegel H, Lea AO 1975. Relationship between protein and proteolytic activity in the midgut of mosquitoes. J Ins Physiol 21: 1597-1604. Bruce MC, Alano P, Duthie S, Carter R 1990. Commitment of the malaria parasite Plasmodium falciparum to sexual and asexual development. Parasitology 100: 191- 200. Canning EU, Sinden RE 1973. The organisation of the ookinete and observations on nuclear division in oocysts of Plasmodium berghei. Parasitology 67: 29- 40. Carter R, Bushell G, Saul A, Graves PM, Kidson C 1985. Two apparently nonrepeated epitopes on gametes of Plasmodium falciparum are targets of transmission-blocking antibodies. Infect Immun 50: 102-106. Carter R, Chen DH 1976. Malaria transmission blocked by immunisation with gamets of the malaria parasite. Nature 263: 57-60. Carter R, Graves PM 1988. Gametocytes. In: Malaria. Principles and practice of malariology, edited by Wernsdorfer, W.H. and McGregor, Sir I. Edinburgh: Chuchill Livingstone, p. 253- 306. Carter R, Graves PM, Creasey A, Byrne K, Read D, Alano P, Fenton B 1989a. Plasmodium falciparum: an abundant stage-specific protein expressed during early gametocyte development. Exp Parasitol 69: 140-149. Carter R, Graves PM, Keister DB, Quakyi IA 1990. Properties of epitopes of Pfs 48/45, a target of transmission blocking monoclonal antibodies, on gametes of different isolates of Plasmodium falciparum. Paras Immunol 12: 587-603. Carter R, Graves P M, Quakyi I, Good MF 1989b. Restricted or absent immune responses in human populations to Plasmodium falciparum gamete antigens that are targets of malaria transmission-blocking antibodies. J Exp Med 169: 135-147. Carter R, Gwadz RW, Mcauliffe FM 1979. Plasmodium gallinaceum: transmission blocking immunity in chickens. I. Comparative immunogenicity of gametocyte and gamete- containing preparations. Exp Parasitol 47: 185-193. Carter R, Kaushal DC 1984. Characterization of antigens on mosquito midgut stages of Plasmodium gallinaceum. III. changes in zygote surface proteins during transformation to mature ookinete. Mol Biochem Parasitol 13: 235- 241. Carter R, Kumar N, Quakyi I, Good M, Mendis K, Graves P, Miller LH 1988. Immunity to sexual stages of malaria parasites. Progr Allergy 41: 193-214. Carter R, Nijhout MM 1977. Control of gamete formation (exflagellation) in malaria parasites. Science 195: 407-409. Cattani JA 1989. Malaria vaccines: results of human trials and directions of current research. Exp Med 68: 242- 247. Cerami C, Frevert V, Sinnis P, Takacs B, Clavijo P, Santos MJ, Nussenzweig V 1992b. The basolateral domain of the hepatocyte plasma membrane bears receptors for the CSP of Plasmodium falciparum sporozoites. Cell 70: 1021- 1033.
Cerami C, Kwakyeberko F, Nusswenzweig V 1992a. Binding of
malarial circumsporozoite protein to sulfatides
Chagnon A, Yao N, Carli P, Paris JF, Marlier S, Pierre C,
Bassiere H 1992. La proteine C-reactive dans l'acces palustre.
Presse Med 21: 217-218.
Charoenvit Y, Leef MF, Yuan LF, Sedegah M, Beaudoin RL 1987.
Characterization of Plasmodium yoelii monoclonal
antibodies directed against stage-specific sporozoite
antigens. Infect Immun 55: 604-608.
Chege GMM, Pumpuni CB, Beier JC 1996. Proteolytic enzyme
activity and Plasmodium falciparum sporogonic
development in three species of Anopheles mosquitoes.
J Parasitol 82: 11-16.
Chutmongkonkul M, Maier WA, Seitz HM 1992. Plasmodium
falciparum: effect of chloroquine, halofantrine and
pyrimethamine on the infectivity of gametocytes for An.
stephensi mosquitoes. Ann Trop Med Parasitol
86: 103-110.
Coleman RE, Vaughan JA, Hayes DO, Hollingdale MR, Do Rosario
VE 1988. Effect of mefloquine and artisinin on the sporogonic
cycle of Plasmodium berghei ANKA in Anopheles
stephensi mosquitoes. Acta Leid 57: 61-
74.
Collins FH 1994. Prospects for malaria control through genetic
manipulation of its vectors. Parasitol Today 10:
370-371.
Collins FH, Sakai RK, Vernick KD, Paskewitz S, Seeley DC,
Miller LH, Collins WE, Campbell CC, Gwadz RW 1986. Genetic
selection of a Plasmodium refractory strain of the
malaria vector Anopheles gambiae. Science
234: 607-610.
Coluzzi M 1992. Malaria vector analysis and control.
Parasitol Today 8: 113-118.
Cornelissen AWCA 1988. Sex determination and sex
differentiation in malaria parasites. Biol Rev
63: 379-394.
Crampton JM 1994. Molecular studies of insect vectors of
malaria. Adv Parasitol 34: 1-31.
Crampton JM, Galler R, Sinden RE, Crisanti A 1993. La lutte
genetique contre les moustiques. La Recherche
259: 1218-1219.
Crampton JM, Morris A, Lycett G, Warren A, Eggleston P 1990.
Transgenic mosquitoes: a future vector control strategy?
Parasitol Today 6: 31-36.
Curtis CF 1994. The case for malaria cntrol by genetic
manipulation of its vectors. Parasitol Today 10:
371-374.
Daher VR, Krettli AU 1980. Infectivity of Plasmodium
gallinaceum sporozoites from oocysts. J Protozool
27: 440-442.
Davies EE 1974. Ultrastructural studies on the early ookinete
stage of Plasmodium berghei nigeriensis and its
transformation into an oocyst. Ann Trop Med Parasitol
68: 283-290.
Dearsly AL, Sinden RE, Self IA 1990. Sexual development in
malarial parasites - Gametocyte production, fertility and
infectivity to the mosquito vector. Parasitology
100: 359-368.
De Zoysa APK, Herath PRJ, Abhywrana A, Padmalal UKGK, Mendis
KN 1988. Modulation of human malaria transmission by anti-
gamete transmission blocking immmunity. Trans R Soc Trop
Med Hyg 82: 548-553.
Do Rosario .E, Vaughan JA, Murphy M, Harrod V, Coleman R 1988.
Effect of chloroquine on the sporogonic cycle of Plasmodium
falciparum and Plasmodium berghei in anopheline
mosquitoes. Acta Leid 57: 53-60.
Duffy P, Pimenta P, Kaslow DC 1993. Pgs28 belongs to a family
of epidermal growth factor-like antigens that are targets of
malaria transmission-blocking antibodies. J Exp Med
177: 505-510.
Duncan D, Eades J, Julian SR, Micks D 1960. Electron
microscopic observations on malarial oocysts (Plasmodium
relictum). J Protozool 7: 18-26.
Eyles DE 1952. Studies on Plasmodium gallinaceum II.
Factors in the blood of the vertebrate host influencing
mosquito infection. Am J Trop Med Hyg 5: 276-
290.
Feldmann AM, Billingsley PF, Savelkoul A 1990. Blood meal
digestion by strains of Anopheles stephensi Liston
(Diptera: Culicidae) of differing susceptibility to
Plasmodium falciparum. Parasitology 101:
193-200.
Feldmann AM, Ponnudurai T 1989. Selection of Anopheles
stephensi for refractoriness and susceptibility to
Plasmodium falciparum. Med Vet Entomol
3: 41-52.
Foo A, Carter R, Lambros C, Graves P, Quakyi I, Targett GAT,
Ponnudurai T, Lewis GE 1991. Conserved and variant epitopes of
target antigens of transmission-blocking antibodies among
isolates of Plasmodium falciparum from Malaysia. Am
J Trop Med Hyg 44: 623-631.
Fowler RE, Billingsley PF, Pudney M, Sinden RE 1994.
Inhibitoy action of the anti-malarial Atovaquone (566C80)
against Plasmodium berghei ANKA in the mosquito Anopheles
stephensi. Parasitology 108: 383-388.
Fowler RE, Sinden RE, Pudney M 1995. Inhibitory activity of
the anti-malarial Atovaquone (566C80) against ookinete,
oocysts and sporozoites of Plasmodium berghei. J
Parasitology 8: 452-458.
Freier J E, Friedman S 1987. Effect of Plasmodium
gallinaceum infection on the mortality and body weight of
Aedes aegypti (Diptera : Culicidae). J Med
Entomol 24: 6-10.
Freyvogel TA 1966. Shape, movement in situ and locomotion of
plasmodial ookinetes. Acta Trop 23: 201-221.
Fries HCW, Lamers MBAC, Smits MA, Ponnudurai T, Meuwissen JHET
1989. Characterization of epitopes on the 25 kiloDalton
protein of the macrogametes/zygotes of Plasmodium
falciparum. Parasite Immunol 11: 31-45.
Gamage-Mendis AC, Rajakaruna J, Carter R, Mendis KN 1992.
Transmission blocking immunity to human Plasmodium
vivax malaria in an endemic population in Kataragama, Sri-
Lanka. Parasite Immunol 14: 385-396.
Gamage-Mendis AC, Rajakaruna J, Weerasinghe S, Mendis C,
Carter R, Mendis KN 1993. Infectivity of Plasmodium
vivax and Plasmodium falciparum to Anopheles
tessellatus; relationship between oocyst and sporozoite
development. Trans R Soc Trop Med Hyg 87: 3-
6.
Garnham PCC 1966. Malaria parasites and other
haemosporidia. Blackwell Scientific Publications, Oxford,
1114 pp.
Garnham PCC, Bird RG, Baker JR 1962. Electron microscopic
studies of motile stages of malarial parasites III. The
ookinetes of Haemamoeba and Plasmodium. Trans
R Soc Trop Med Hyg 56: 116-120.
Garnhm PCC, Bird RG, Baker JR, Desser SS, El-Nahal HMS 1969.
Electron microscopic studies on the motile stages of malaria
parasites VI. The ookinete of Plasmodium berghei yoelii
and its transformation into an early oocyst. Trans R Soc
Trop Med Hyg 63: 187-194.
Gass RF 1977. Influences of blood digestion on the development
of Plasmodium gallinaceum (Brumpt) in the midgut of
Aedes aegypti (L.). Acta Trop 34: 127-
140.
Gass RF, Yeates RA 1979. In vitro damage to cultured
ookinetes of Plasmodium gallinaceum by digestive
proteinases from susceptible Aedes aegypti. Acta
Trop 36: 243-252.
Golenda CF, Starkweather WH, Wirtz RA 1990. The distribution
of circumsporozoite protein (Cs) in Anopheles stephensi
mosquitoes infected with Plasmodium falciparum malaria.
J Histochem Cytochem 38: 475-481.
Good MF 1991a. Towards the development of the ideal malaria
vaccine - A decade of progress in a difficult field. Med
J Austr 154: 284-289.
Good MF 1991b. The implications for malaria vaccine programs
if memory T-cells from non-exposed humans can respond to
malaria antigens. Curr Op Immunol 3: 496-
502.
Good MF 1992. A malaria vaccine strategy based on the
induction of cellular immunity. Immunol Today 13: 126-
129.
Goonewardene R, Carter R, Gamage CP, Delgiudice G, David PH,
Howie S, Mendis KN 1990. Human T cell proliferative responses
to Plasmodium vivax antigens: evidence of
immunosuppression following prolonged exposure to endemic
malaria. Europ J Immunol 20: 1387-1391.
Gordon DM, Davis DR, Lee M, Lambros C, Harrison BA, Samuel R,
Campbell GH, Jeghathesan M, Selvarajan K, Lewis GE 1991.
Significance of circumsporozoite-specific antibody in the
natural transmission of Plasmodium falciparum, Plasmodium
vivax, and Plasmodium malariae in an aboriginal
(Orang-Asli) population of central peninsular Malaysia. Am
J Trop Med Hyg 45: 49-56.
Graf R, Briegel H 1982. Comparison between aminopeptidase and
trypsin activity in blood-fed females of Aedes
aegypti. Rev Suis Zool 89: 845-850.
Graf R, Raikhel AS, Brown MR, Lea AO, Briegel H 1986. Mosquito
trypsin: immunocytochemical localization in the midgut of
blood-fed Aedes aegypti (L.). Cell Tis Res
245: 19-27.
Grau GE, Taylor TE, Molyneux ME, Wirima JJ, Vassali P, Hommel
M, Lambert PH 1989. Tumor necrosis factor and disease severity
in children with falciparum malaria. N Engl J Med
320: 1586-1591.
Graves PM, Carter R, Burkot TR, Quakyi IA, Kumar N 1988.
Antibodies to Plasmodium falciparum gamete surface
antigens in Papua New Guinea sera. Paras Immunol 10:
209-218.
Graves PM, Doubrovsky A, Satabongkot J, Battistutta D 1992.
Human antibody responses to epitopes on the Plasmodium
falciparum gametocyte antigen PFS48/45 and their
relationship to infectivity of gametocyte carriers. Am J
Trop Med Hyg 46: 711-719.
Grotendorst CA, Carter, R 1987. Complement effects on the
infectivity of Plasmodium gallinaceum to Aedes
aegypti mosquitoes II. changes in sensitivity to
complement-like factors during zygote development. J
Parasitol 73: 980-984.
Grotendorst CA, Carter R, Rosenberg R, Koontz L 1986.
Complement effects on the infectivity of Plasmodium
gallinaceum to Aedes aegypti mosquitoes I.
Resistance of zygotes to the alternative pathway of
complement. J Immunol 136: 4270-4274.
Grotendorst CA, Kumar N, Carter R, Kaushal DC 1984. A surface
protein expressed during transformation of zygotes of
Plasmodium gallinaceum is a target of transmission-
blocking antibodies. Infec Immun 45: 775-777.
Gwadz W 1976. Malaria: successful immunization against the
sexual stages of Plasmodium gallinaceum. Science
193: 1150-1151.
Gwadz RW, Kaslow D, Lee J-Y, Maloy WL, Zasloff M, Miller LH
1989. Effects of magainins and cercropins on the sporogonic
development of malaria parasites in mosquitoes. Infec
Immun 57: 2628-2633.
Hamilton AJ, Davies CS, Sinden RE 1988. Expression of the
circumsporozoite proteins revealed in situ in the mosquito
stages of Plasmodium berghei by the Lowicryl-immunogold
technique. Parasitology 96: 273-280.
Harte PG, Rogers N, Targett GAT 1985a. Vaccination with
purified microgamete antigens prevents transmission of rodent
malaria. Nature 316: 258-259.
Harte PG, Rogers NC, Targett GAT 1985b. Monoclonal anti-
gamete antibodies prevent transmission of murine malaria.
Paras Immunol 7: 607-615.
Hawking F, Wilson ME, Gammage K 1971. Evidence for the cyclic
development and short-lived maturity in the gametocytes of
Plasmodium falciparum. Trans R Soc Trop Med
Hyg 65: 549-559.
Hecker H 1977. Structure and function of midgut epithelial
cells in Culicidae mosquitoes (Insecta, Diptera).
Cell Tis Res 184: 321-341.
Hollingdale MR, do Rosario V 1989. Malaria transmission-
enhancing activity in mosquitoes by mammalaian host anti-
sporozoite antibodies. Exp Parasitol 68: 365-
368.
Hommel M 1991. Steps towards a malaria vaccine. Res
Immunol 142: 618-631.
Houk EJ, Hardy JL, Chiles RE 1986. Mesenteronal epithelial
cellsurface charge of the mosquito, Culex
tarsalis Coquillett. Binding of colloidal iron
hydroxide, native ferritin and cationised ferritin. J
Submicroscop Cytol 18: 385-396.
Howells RE, Davies EE 1971. Nuclear division in the oocyst of
Plasmodium berghei. Ann Trop Med Hyg 65:
451-459.
Huber M, Cabib E, Miller LH 1991. Malaria parasite chitinase
and penetration of the mosquito peritrophic membrane. Proc
Natl Acad Sci USA 88: 2807-2810.
Ichimori K, Curtis CF, Targett GAT 1990. The effects of
chloroquine on the infectivity of chloroquine-sensitive and -
resistant populations of Plasmodium yoelii nigeriensis
to mosquitoes. Parasitology 100: 377-381.
Iinselburg J 1983. Gametocyte formation by the progeny of
single Plasmodium falciparum schizonts. J
Parasitol 69: 584-591.
James AA, Rossignol PA 1991. Mosquito salivary glands -
Parasitological and molecular aspects. Parasitol Today
7: 267-271.
Jeffery GM, Eyles DE 1955. Infectivity to mosquitoes of
Plasmodium falciparum as related to gametocyte density
and duration of infection. Am J Trop Med Hyg 4:
781-789.
Jeffery GM, Young MD, Eyles DE 1956. The treatment of
Plasmodium falciparum infection with chloroquine, with
a note on infectivity to mosquitoes of primaquine-and
pyrimethamine-treated cases. Am J Hyg 64: 1-
11.
Karunaweera ND, Grau GE, Gamage P, Carter R, Mendis KN 1992.
Dynamics of fever and serum levels of tumor necrosis factor
are closely associated during clinical paroxysms in
Plasmodium vivax malaria. Proc Natl Acad Sci USA
89: 3200-3203.
Kaslow DC, Bathurst IC, Barr PJ 1992. Malaria trasmission-
blocking vaccines. Trends Biotechnol 10: 388-
391.
Kaslow DC, Bathurst IC, Lensen T, Ponnudurai T, Barr PJ,
Keister DB 1994. Saccharomyces cerevisiae recombinant Pfs25
adsorbed to alum elecits antibodies that block transmission of
Plasmodium falciparum. Infec Immun 62: 5576-5580.
Kaslow DC, Isaacs SN, Quakyi IA, Gwadz RW, Moss B, Keister DB
1991. Induction of Plasmodium falciparum transmission
blocking antibodies by recombinant vaccinia virus.
Science 252: 1310-1313.
Kaslow DC, Nussenzweig V, Miller L 1994a. Meeting on parasites
and the invertebrate vector.
Mem Inst Oswaldo
Cruz 89: 279-295.
Kaslow DC, Quakyi IA, Syin C, Raum MG, Keister DB, Coligan
JE, McCutchan TF, Miller LH 1988. A vaccine candidate from
the sexual stage of human malaria that contains EGF-like
domains. Nature 333: 74-76.
Kaushal DC, Carter R 1984. Caracterization of antigens on the
mosquito midgut stages of Plasmodium gallinaceum . II.
Comparison of surface antigens of male and female gametes and
zygotes. Mol Biochem Parasitol 11: 145-156.
Kaushal DC, Carter R, Howard RJ, Macauliffe FM 1983a.
Characterization of antigens on mosquito midgut stages of
Plasmodium gallinaceum I. Zygote surface antigens.
Mol Biochem Parasitol 8: 53-69.
Kaushal DC, Carter R, Rener J, Grotendorst CA, Miller LH,
Howard RJ 1983b. Monoclonal antibodies against surface
determinants on gametes of Plasmodium gallinaceum block
transmission of malaria prasites to mosquitoes. J
Immunol 131: 2557-2562.
Kawamoto F, Alejo Blanco R, Fleck SL, Sinden RE 1991.
Plasmodium berghei - Ionic regulation and the induction
of gametogenesis. Exp Parasitol 72: 33-42.
Kern P, Hemmer CJ, Van Damme J, Gruss HJ, Dietrich M 1989.
Elevated tumor necrosis factor-alfa and interleukin-6 serum
levels as markers for complicated Plasmodium falciparum
malaria. Am J Med 87: 139-143.
Kidwell MG, Ribeiro JMC 1992. Can transposable elements be
used to drive disease refractoriness genes into vector
populations? Parasitol Today 8: 325-329.
King C 1988. Cell motility of sporozoan protozoa.
Parasitol Today 4: 315-319.
Kitthawee S, Edman JD, Sattabongkot J 1990. Evaluation of
survival potential and malaria susceptibility among different
size classes of laboratory-reared Anopheles dirus.
Am J Trop Med Hyg 43: 328-332.
Kumar N, Carter R 1984. Biosynthesis of the target antigens of
antibodies blocking transmission of Plasmodium
falciparum. Mol Biochem Parasitol 13: 333-
342.
Kumar N, Carter R 1985. Biosynthesis of two stage-specific
membrane proteins during transformation of Plasmodium
gallinaceum zygotes into ookinetes. Mol Biochem
Parasitol 14: 127-139.
Kwiatowski D 1992. Malaria: becoming more specific about non-
specific immunity. Curr Op Immunol 4: 425-
431.
Kwiatowski D, Hill AVS, Sambou I, Twumasi P, Castracane J,
Manogue KR, Cerami A, Brewster DR, Greenwood BM 1990. TNF
concentrations in fatal cerebral, non-fatal cerebral, and
uncomplicated Plasmodium falciparum malaria.
Lancet ii: 1201-1204.
Lensen AHW, van Gemert GJA, Bolmer MG, Meis JFGM, Kaslow D,
Meuwissen JHE Th, Ponnudurai T 1993. Transmission blocking
antibody of the Plasmodium falciparum zygote/ookinete
surface protein Pfs25 also influences sporozoite development.
Parasite Immunol 14: 471-479.
Li X, Sina B, Rossignol PA 1992. Probing behaviour and
sporozoite delivery by Anopheles stephensi infected
with Plasmodium berghei. Med Vet Entomol 6:
57-61.
Lines J.D, Wilkes TJ, Lyimo EO 1991. Human malaria
infectiousness measured by age-specific sporozoite rates in
An gambiae in Tanzania. Parasitology 102:
167-177.
Lombardi S, Esposito F, Zavala F, Lamizana L, Rossi P,
Sabatinelli G, Nussenzweig R, Coluzzi M 1987. Detection and
anatomical localization of Plasmodium falciparum
circumsporozoite protein and sporozoites in the afrotropical
malaria vector Anopheles gambiae S.L. Am J Trop Med
Hyg 37: 491-494.
Lyimo EO, Koella JC 1992. Relatiosnship between body size of
adult Anopheles gambiae s.l. and infection with the
malaria parasite Plasmodium falciparum.
Parasitology 104: 233-237.
Mack SR, Samuels S, Vanderberg JP 1979. Hemolymph of
Anopheles stephensi from uninfected and Plasmodium
berghei-infected mosquitoes. 2. Free amino acids. J
Parasitol 65: 130-136.
Mack SR, Vanderberg JP 1978. Hemolymph of Anopheles
stephensi from noninfected and Plasmodium berghei-
infected mosquitoes. 1. Collection procedure and physical
characteristics. J Parasitol 64: 918-923.
Maier WA, Becker-Feldman H, Seitz HM 1987. Pathology of
malaria infected mosquitoes. Parasitol Today 3:
216-218.
Martin SK, Miller LH, Nijhout MM, Carter R 1978. Plasmodium
gallinaceum: induction of male gametocyte exflagellation
by
phosphdiesterase inhibitors. Exp Parasitol 44:
239-242.
Mehlhorn H, Peters W, Haberkorn A 1980. The formation of
ookinetes and oocysts in Plasmodium gallinaceum
(Haemosporidia) and considerations on phylogenetic
relationships between Haemosporidia, Piroplasmida and other
Coccidia. Protistologica 16: 135-154.
Meis JFGM, Croes H, Mons B, Vanbelkum A, Ponnudurai T 1992a.
Localization of circumsporo-zoite protein in the sporogonic
stages of Plasmodium vivax. Parasitol Res
78: 165-167.
Meis JFGM, Ponnudurai T 1987. Ultrastructural studies on the
interaction of Plasmodium falciparum ookinetes with the
midgut epithelium of Anopheles stephensi mosquitoes.
Parasitol Res
73: 500-506.
Meis JFGM, Pool G, Vangemert GJ, Lensen AHW, Ponnudurai T,
Meuwissen JHET 1989. Plasmodium falciparum ookinetes
migrate intercellularly through Anopheles stephensi
midgut epithelium. Parasitol Res 76: 13-19.
Meis JFGM, Verhave JP 1988. Exoerythrocytic development of
malarial parasites. Adv Parasitol 27: 1-66.
Meis JFGM, Wismans PGP, Jap PHK, Lensen AHW, Ponnudurai T
1992b. A scanning electron microscopic study of the sporogonic
development of Plasmodium falciparum in Anopheles
stephensi. Acta Trop 50: 227-236.
Mendis KN, Carter R 1991. Transmission blocking immunity may
provide clues that antimalarial immunity is largely T-
independent. Res Immunol 142: 687-690.
Mendis KN, David PH, Carter R 1990b. Human immune responses
against sexual stages of malaria parasites: considerations for
malaria vaccines. Intern J Parasitol 20: 497-
502.
Mendis, KN, Ezoysa APK, Abhayawardena TA, Carter R, Herath
PRJ, Mendis G 1990a. Characteristics of malaria transmission
in Kataragama, Sri-Lanka - A focus for immuno-epidemiological
studies. Am J Trop Med Hyg 42: 298-308.
Mendis KN, Munesinghe YD, DeSilva YNY, Keragalla I, Carter R
1987. Malaria transmission-blocking immunity induced by
natural infections of Plasmodium vivax in humans.
Infec Immun 55: 369-372.
Mendis KN, Naotunne TD, Karunaweera ND, Delgiudice G, Grau GE,
Carter R 1990c. Anti-parasite effects of cytokines in malaria.
Immunol Lett 25: 217-220.
Mitchell GH 1989. An update on candidate malaria vaccines.
Parasitology 98: S29-S47.
Mons B 1985. Induction of sexual differentiation in malaria.
Parasitol Today 1: 87-89.
Mons B 1986. Intraerythrocytic differentiation of
Plasmodium berghei. Acta Leid 54: 1-
83.
Mons B, Janse CJ, Boorsma EG 1985. Synchronized erythrocytic
schizogony and gametocytogenesis of Plasmodium berghei
in vivo and in vitro. Parasitology 91: 423-430.
Muirhead-Thomson RC 1957. The malarial infectivity of an
African village population to mosquitoes (Anopheles
gambiae). A random xenodiagnostic survey. Am J Trop
Med Hyg 6: 971-979.
Nagasawa H, Aikawa M, Procell PM, Campbell GH, Collins WE,
Campbell CC 1988. Plasmodium malariae: distribution of
circumsporozoite protein in the oocyst and salivary gland
sporozoite. Exp Parasitol 66: 27-34.
Nagasawa H, Procell PM, Atkinson CT, Campbell GH, Collins WE,
Aikawa M 1987. Localization of circumsporozoite protein of
Plasmodium ovale in midgut oocysts. Infec Immun
55: 2928-2932.
Naik P, Voller A 1984. Serum C-reactive protein levels and
falciparum malaria. Trans R Soc Trop Med Hyg 78: 812-
813.
Naotunne TD, Karunaweera ND, Delgiudice G, Kularatne MU, Grau
GE, Carter R, Mendis KN 1991. Cytokines kill malaria
parasites during infection crisis - Extracellular
complementary factors are essential. J Exp Med
173: 523-529.
Naotunne TD, Rathnayake KDL, Jayasinghe A, Carter R, Mendis
KN 1990. Plasmodium cynomolgi - Serum-mediated blocking
and enhancement of infectivity to mosquitoes during infections
in the natural host, Macaca sinica. Exp
Parasitol 71: 305-313.
Nijhout MM 1979. Plasmodium gallinaceum: exflagellation
stimulated by a mosquito factor. Exp Parasitol
48: 75-80.
Nijhout MM, Carter R 1978. Gamete development in malarial
parasites: bicarbonate-dependent stimulation by pH in vitro.
Parasitology 76: 39-53.
Nussenzweig RS 1990. Recombinant proteins and synthetic
peptides as malaria vaccine candidates. Can J Microbiol
36: 817-820.
Nussenzweig V, Nussenzweig RS 1989. Rationale for the
development of an engineered sporozoite malaria vaccine.
Adv Immunol 45: 283-334.
Ono T, Onhnishe Y, Nagamune K, Kano M 1993. Gametocytogenesis
induction by Berenil in cultured Plsmodium falciparum.
Exp Parasitol 77: 74-78.
Pancake SJ, Holt GD, Mellouk S, Hoffman SL 1992. Malaria
sporozoites and circumsporozoite proteins bind specifically to
sulfated glycoconjugates. J Cell Biol 117:
1351-1357.
Paskewitz SM, Brown MR, Collins FH, Lea AO 1989.
Ultrastructural localization of phenoloxidase in the midgut of
refractory Anopheles gambiae and association of the
enzyme with encapsulated Plasmodium cynomolgi. J
Parasitol 75: 594-600.
Paton MG, Barker GC, Matsuoka H, Ramesar J, Janse CJ, Waters
AP, Sinden RE 1993. Structure and expression of a post-
transcriptionally regulated malaria gene enconding a surface
protein from the sexual stages of Plasmodium berghei.
Mol Biochem Parasitol 59: 263-276.
Peiris JSM, Premawansa S, Ranawaka MBR, Udagama PV, Munasonghe
YD, Nanayakkara MV, Gamage CP, Carter R, David PH, Mendis KN
1988. Monoclonal and polyclonal antibodies both block and
enhance transmission of human Plasmodium vivax malaria.
Am J Trop Med Hyg 39: 26-32.
Perrone JB, DeMaio J, Spielman A 1986. Regions of mosquito
salivary glands distinguished by surface lectin-binding
characteristics. Insect Biochem 16: 313-318.
Perrone JB, Spielman A 1988. Time and site of assembly of the
peritrophic membrane of the mosquito Aedes aegypti.
Cell Tiss Res 252: 473-478.
Phillips RS 1992. Vaccination against malaria.
Immunobiology 184: 240-262.
Ponnudurai T, Billingsley PF, Rudin W 1988. Differential
infectivity of Plasmodium for mosquitoes. Parasitol
Today 4: 319-321.
Ponnudurai T, Lensen AHW, Van Gemert, GJA, Bensink MPE, Bolmer
M, Meuwissen JHET 1989. Infectivity of cultured Plasmodium
falciparum gametocytes to mosquitoes. Parasitology
98: 165-173.
Ponnudurai T, Lensen AHW, Van Gemert GJA, Bolmer MG, Meuwissen
JHET 1991. Feeding behaviour and sporozoite ejection by
infected Anopheles stephensi. Trans R Soc Trop Med
Hyg 85: 175-180.
Posthuma G, Meis FGM, Verhave JP, Hollingdale MR, Ponnudurai
T, Meuwissen JHET 1988. Localization of circumsporozoite
protein of the malaria parasite Plasmodium falciparum
during sporogony in Anopheles stephensi midguts using
immunoelectron-microscopy of ultrathin cryosections. Eur J
Cell Biol 46: 18-24.
Posthuma G, Meis JFGM, Verhave JP, Gigengak S, Hollingdale MR,
Ponnudurai T, Genze HJ 1989. Immunogold determination of
Plasmodium falciparum circumsporozoite protein in
Anopheles stephensi salivary gland cells. Eur J
Cell Biol 49: 66-72.
Premawansa S, Gamage-Mendis A, Perera L, Begarnie S, Mendis K,
Carter R 1994. Plasmodium falciparum malaria
trasmission-blocking immunity under conditions of low
endemicity as in Sri Lanka. Parasite Immunol 16:
35-42.
Premawansa S, Peiris JSM, Perera KLRL, Ariyaratne G, Carter R,
Mendis KN 1990. Target antigens of transmission blocking
immunity of Plasmodium vivax malaria - Characterization
and polymorphism in natural parasite isolates. J
Immunol 144: 4376-4383.
Pringle G 1965. A count of the sporozoites in an oocyst of
Plasmodium falciparum. Trans R Soc Trop Med Hyg
59: 289-290.
Pringle G 1966. A quantitative study of naturally acquired
malaria infections in Anopheles gambiae and
Anopheles funestus in a highly malarious area of east
Africa. Trans R Soc Trop Med Hyg 60: 626-
632.
Quakyi IA, Carter R, Rener J, Kumar N, Good MF, Miller LH
1987. The 230-kDa gamete surface protein of Plasmodium
falciparum is also a target for transmission-blocking
antibodies. J Immunol 139: 4213-4217
Ramasamy M S, Ramasamy R 1990. Effect of anti-mosquito
antibodies on the infectivity of the rodent malaria parasite
Plasmodium berghei to Anopheles farauti. Med
Vet Entomol 4: 161-166.
Ranawaka G, Alejo Blanco R, Sinden RE 1993. The effect of
transmission-blocking antibody ingested in primary and
secondary bloodfeeds, upon the development of Plasmodium
berghei in the mosquito vector. Parasitology
107: 225-231.
Ranawaka GRR, Fleck SL, Alejo Blanco AR, Sinden RE 1994.
Characterization of the modes of action of anti-Pbs21 malaria
transmission-blocking immunity: ookinete to oocyst
differentiation in vivo. Parasitology
109: 403-411.
Ranawaka MB, Munesinghe YD, De Silva DMR, Carter R, Mendis KN
1988. Boosting of transmission-blocking immunity during
natural Plasmodium vivax infections in humans depends
upon frequent reinfection. Infec Immun 56:
1820-1824.
Ree GH 1971. C-reactive protein in Gambian africans with
special reference to P. falciparum malaria. Trans R.
Soc Trop Med Hyg 65: 574-580.
Rener J, Graves PM, Carter R, Williams JL, Burkot T 1983.
Target antigens of transmission-blocking immunity on gametes
of Plasmodium falciparum. J Exp Med 158:
971-976.
Ribeiro JMC 1987. Role of saliva in blood-feeding by
arthropods. Ann Rev Entomol 32: 463-478.
Ribeiro JMC, Modi GB, Tesh RB 1989. Salivary apirase activity
of some old world phlebotomine sand flies. Insect
Biochem 19: 409-412.
Ribeiro MC, Rossignol PA, Spielman A 1984. Role of mosquito
saliva in blood vessel location. J Exp Biol
108: 1-9.
Ribeiro JMC, Rossignol PA, Spielman A 1985. Salivary gland
apyrase activity determines probing time in Anopheline
mosquitoes. J Insect Physiol 31: 689-692.
Rickman LS, Jones TR, Long GW, Paparello S, Schneider I, Paul
CF, Beaudoin RL, Hoffman SL 1990. Plasmodium
falciparum-infected Anopheles stephensi
inconsistently transmit malaria to humans. Am J Trop Med
Hyg 43: 441-445.
Riley E, Greenwood B 1990. Measuring cellular immune responses
to malaria antigens in endemic populations - Epidemiological,
parasitological and physiological factors which influence in
vitro assays. Immunol Lett 25: 221-229.
Robert V, Verhave JP, Ponnudurai T, Louwe L, Scholtens P,
Carnevale P 1988. Study of the distribution of
circumsporozoite antigen in Anopheles gambiae infected
with Plasmodium falciparum, using the enzyme-linked
immunosorbent assay. Trans R Soc Trop Med Hyg
82: 389-391.
Robson KJH, Hall JRS, Jennings MW, Harris TJR, Marsh K,
Newbold CI, Tate VE, Weatherhall DJ 1988. A highly conserved
amino-acid sequence in thrombospondin, properdin and in
proteins from sporozoites and blood stages of a human malaria
parasite. Nature 335: 79-82.
Rogers WO, Malik A, Mellouk S, Nakamura K, Rogers MD, Szarfman
A, Gordon DM, Nussler AK, Aikawa M, Hoffman SL 1992b.
Characterization of Plasmodium falciparum sporozoite
surface protein 2. Proc Natl Acad Sci USA 89: 9176-
9180.
Rogers WO, Rogers MD, Hedstrom RC, Hoffman SL 1992a.
Characterization of the gene encoding sporozoite surface
protein-2, a protective Plasmodium yoelii sporozoite
antigen. Mol Biochem Parasitol 53: 45-51.
Rosenberg R 1985. Inability of Plasmodium knowlesi
sporozoites to invade Anopheles freeborni salivary
glands. Am J Trop Med Hyg 34: 687-691.
Rosenberg R, Koontz LC 1984. Plasmodium gallinaceum:
density dependent limits on infectivity to Aedes
aegypti. Exp Parasitol 57: 234-238.
Rosenberg R, Koontz LC, Alston K, Friedman FK 1984.
Plasmodium gallinaceum: erythrocyte factor essential
for zygote infection of Aedes aegypti. Exp
Parasitol 57: 158-164.
Rosenberg R, Rungsiwongse J 1991. The number of sporozoites
produced by individual malaria oocysts. Am J Trop Med
Hyg 45: 574-577.
Rosenberg R, Wirtz RA, Scneider I, Burge R 1990. An
estimation of the number of malaria sporozoites ejected by a
feeding mosquito. Trans R Soc Trop Med Hyg 84:
209-212.
Rossignol PA, Ribeiro JMC, Jungery M, Turell MJ, Spielman A,
Bailey CL 1985. Enhanced mosquito blood-finding success on
parasitaemic hosts: evidence for vector-parasite mutualism.
Proc Natl Acad Sci USA 82: 7725-7727.
Rossignol PA, Ribeiro JMC, Spielman A 1984. Increased
intradermal probing time in sporozoite-infected mosquitoes.
Am J Trop Med Hyg 33: 17-20.
Rossignol PA, Ribeiro JMC, Spielman A 1986. Increased biting
rate and reduced fertility in sporozoite-infected mosquitoes.
Am J Trop Med Hyg 35: 277-279.
Roush RT 1993. Occurrence, genetics and management of
insecticide resistance. Parasitol Today 9: 174-
179.
Rowland M, Boersma E 1988. Changes in the spontaneous flight
activity of the mosquito Anopheles stephensi by
parasitization with the rodent malaria Plasmodium
yoelii. Parasitology 97: 221-227.
Rudin W, Billingsley PF, Saladin S 1991. The fate of
Plasmodium gallinaceum in Anopheles stephensi
Liston and possible barriers to transmission. Ann Soc Belg
Med Trop 71: 167-177.
Rudin W, Hecker H 1989. Lectin-binding sites in the midgut of
the mosquitoes Anopheles stephensi Liston and Aedes
aegypti L. (Diptera: Culicidae0. Parasitol Res
75: 268-279.
Rutledge LC, Gould DJ, Tantichareon B 1969. Factors affecting
the infection of anophelines with human malaria in Thailand.
Trans R Soc Trop Med Hyg 63: 613-619.
Santoro F, Cochrane AH, Nussenzweig V, Nardin EH, Nussenzweig
R, Gwadz RW, Ferreira A 1983. Structural similarities among
the protective antigens of spoorzoites from different species
oof malaria parasites. J Biol Chem 258: 3341-
3345.
Sattabongkot J, Manaeechai N, Rosenberg R 1991. Plasmodium
vivax: gametocyte infectivity of naturally infected Thai
adults. Parasitology 102: 27-31.
Schall JJ 1989. The sex ratio of Plasmodium
gametocytes. Parasitology 98: 343-350.
Schapira A, Beales PF, Halloran ME 1993. Malaria: living with
drug resistance. Parasitol Today 9: 168-174.
Scherf A, Behr C, Sarthou JL, Pla M, Rogier C, Trape JF,
Pereira da Silva L, Dubois P 1993. Immune response in mouse
and malaria-exposed humans to peptides derived from Pf11-1, a
highly repetitive megadalton protein of Plasmodium
falciparum. Eur J Immunol 23: 1574-1581.
Scherf A, Petersen C, Carter R, Alano P, Nellson R, Aikawa M,
Mattei D, Pereira da Silava, L. 1992. Characterization of a
Plasmodium falciparum mutant that has deleted the
majority of the gametocyte-specific Pf11-1 locus. Mem Inst
Oswaldo Cruz 87(Suppl. III): 91-94.
Schneweis S, Maier WA, Seitz HM 1991. Haemolysis of infected
erythrocytes - A trigger for formation of Plasmodium
falciparum gametocytes. Parasitol Res 77:
458-460.
Schulman S, Oppenheim JD, Vanderberg JP 1980. Plasmodium
berghei and Plasmodium knowlesi: serum binding to
sporozoites. Exp Parasitol 49: 420-429.
Seitz HM, Maier WA, Rottok M, Becker-Feldman H 1987.
Concomittant infections of Anopheles stephensi with
Plasmodium berghei and Serratia marcesens
additive detrimental effects. Z Bakteriol Mikrobiol
Hyg 266: 155-166.
Shahabuddin M, Toyoshima T, Aikawa M, Kaslow DC 1993.
Transmission-blocking acitivity of a chitinase inhibitor
activation of malarial parasite chitinase by mosquito
protease. Proc Natl Acad Sci USA 90: 4266-
4270.
Shute PG 1945. An investigation into the number of sporozoites
found in the salivary glands of Anopheles mosquitoes.
Trans R Soc Trop Med Hyg 38: 493-498.
Shute PG, Marion M 1951. A study of gametocytes in a West
Africa strain of Plamodium falciparum. Trans R Soc
Trop Med Hyg 44: 421-438.
Shute PG, Maryon M 1954. The effect of pyrimethamine
(Daraprim) on the gametocytes and oocysts of Plasmodium
falciparum and Plasmodium vivax. Trans R Soc
Trop Med Hyg 48: 50-63.
Sieber KP, Huber M, Kaslow D, Banks SM, Torii M, Aikawa M,
Miller LH 1991. The peritrophic membrane as a barrier: its
penetration by Plasmodium gallinaceum and the effect of
a monoclonal antibody to ookinetes. Exp Parasitol
72: 145-156.
Simonetti AB, Billingsley PF, Winger L, Sinden RE 1993.
Kinetics of expression of two major Plasmodium berghei
antigens in the msquito vector Anopheles stephensi.
J Euk Microbiol (J Protozool) 40: 569-576.
Sinden RE 1975. The sporogonic cycle of Plasmodium yoelii
nigeriensis: a scanning electron microscope study.
Protistologica 11: 31-39.
Sinden RE 1983. The cell biology of sexual development in
Plasmodium. Parasitology 86: 7-28.
Sinden RE 1984. The biology of Plasmodium in the
mosquito. Experientia 40: 1330-1343.
Sinden RE 1985. A cell biologist's view of host cell
recognition and invasion by malarial parasites. Trans R
Soc Trop Med Hyg 79: 598-605.
Sinden RE 1978. Cell Biology, p. 85-186. In R Killick-Kendrick
& WA Peters (eds) Rodent Malaria, New York: Academic
Press.
Sinden RE 1989. Recent advances in the parasitology of
malaria. Trans R Soc Trop Med Hyg 83 Suppl:
3-9.
Sinden RE 1991. Asexual blood stages of malaria modulate
gametocyte infectivity to the mosquito vector - possible
implications for control strategies. Parasitology
103: 191-196.
Sinden RE, Butcher GA, Billker O, Fleck SL 1996. Regulation of
infectivity of Plasmodium to the mosquito vector. Adv
Parasitology 38: 54 - 117.
Sinden RE, Croll NA 1975. Citolology and kinetics of
microgametogenesis and fertilization of Plasmodium yoelii
nigeriensis. Parasitology 70: 53-65.
Sinden RE, Garnham PCC 1973. A comparative study on the
ultrastructure of Plasmodium sporozoites within the
oocyst and salivary glands, with particular reference to the
incidence of the micropore. Trans R Soc Trop Med Hyg
67: 631-637.
Sinden RE, Smalley ME 1976. Gametocytes of Plasmodium
falciparum: phagocytosis by leucocytes in vivo and in
vitro. Trans R Soc Trop Med Hyg 70: 344-345.
Sinden RE, Strong K 1978. An ultrastructural study of the
sporogonic development of Plasmodium falciparum in
Anopheles gambiae. Trans R Soc Trop Med Hyg
27: 477-491.
Sinden RE, Winger LA, Hartley RH, Carter HE, Tirawanchai N,
Davies CS, Sluiters JG 1987. Ookinete antigens of
Plasmodium berghei: a light and electron microscopic
immunogold study of the 21kD determinant recognised by
transmission blocking antibodies. Proc R Soc Lond B
230: 443-458.
Sluiters JF, Visser PE, Van Der Kaay, HJ 1986. The
establishment of Plasmodium berghei in mosquitoes of a
refractory and a susceptible line of Anopheles
atroparvus. Z
Parasitenkd 72: 313-322.
Smalley ME 1977. Plasmodium falciparum gametocytes: the
effect of chloroquine on their development. Trans R Soc
Trop Med Hyg 71: 523-529.
Smalley ME, Brown J, Basset NM 1981. The rate of production of
Plasmodium falciparum gametocytes during natural
infections. Trans R Soc Trop Med Hyg 75: 317-
319.
Snewin VA, Premawansa S, Kapilananda GMG, Ratnayaka L, Udagama
PV, Mattei DM, Khouri E, DelGiudice G, Peiris JSM, Mendis KN,
David PH 1995. Transmission blocking immunity in Plasmodium
vivax malaria: antibodies raised against a peptide block
parasite development in the mosquito vector. J Exp Med
181: 357-362.
Snow RW, Schellenberg JRMA, Peshu N, Forster D, Newton CRJC,
Winstanley PA, Mwangi I, Warn PA, Newbold C, Marsh K 1993.
Periodicity and space-time clustering of severe childhood
malaria on the coast of Kenya. Trans R Soc Trop Med Hyg
87: 386-390.
Speer CA, Rosale-Ronquillo MC, Silverman PH 1974. Scanning
electron microscope observations of Plasmodium berghei
ookinetes in primary mosquito cell cultures. J Invert
Pathol 24: 179-183.
Sterling CR, Aikawa M, Vanderberg JP 1973. The passage of
Plasmodium berghei sporozoites through the salivary
glands of Anopheles stephensi: an electron microscope
study. J Parasitol 59: 593-605.
Stewart MJ, Vanderberg JP 1988. Malaria sporozoites leave
behind trails of circumsporozoite protein during gliding.
J Protozool 35:389-393.
Stewart MJ, Vanderberg JP 1991. Malaria sporozoites release
circumsporozoite protein from their apical end and translocate
it along their surface. J Protozool 38: 411-
421.
Stewart MJ, Vanderberg JP 1992. Electron microscopic analysis
of circumsporozoite protein trail formation by gliding malaria
sporozoites. J Protozool 39: 663-671.
Stohler H 1957. Analyse des infektionsverlaufes von
Plasmodium gallinaceum im darme von Aedes
aegypti. Acta Trop 14: 302-352.
Strome CPA, Beaudoin RL 1974. The surface of the malarial
parasite I. Scanning electron microscopy of the oocyst.
Exp Parasitol 36: 131-142.
Syafruddin A, Arakawa R, Kamimura, Kawamoto F 1991.
Penetration of the mosquito midgut wall by the ookinetes of
Plasmodium yoelii nigeriensis. Parasitol Res
77: 230-236.
Syafruddin A, Arakawa R, Kamimura K, Kawamotoo F 1992.
Development of Plasmodium berghei ookinetes to young
oocysts in vitro. J Protozool 39: 333-338.
Targett GAT 1989. Status of malaria vaccine research. J R
Soc Med 82: 52-56.
Targett GAT 1990. Immunity to sexual stages of human malaria
parasites-Immune modulation during natural infections,
antigenic determinants, and the induction of transmission-
blocking immunity. Scand J Infec Dis 76: 79-88.
Terzakis JA 1971. Transformation of the Plasmodium
cynomolgi oocyst. J Protozool 18: 62-73.
Tirawanchai N, Winger LA, Nicholas J, Ssinden RE 1991.
Analysis of immunity induced by the affinity-purified 21-
Kilodalton zygote-ookinete surface antigen of Plasmodium
berghei. Infec Immun 59: 36-44.
Titus RG, Ribeiro JMC 1990. The role of vector saliva in
transmission of arthropod-borne disease. Parasitol
Today 6: 157-160.
Torii M, Nakamura K, Sieber KP, Miller LH, Aikawa M 1992a.
Penetration of the mosquito (Aedes aegypti) midgut wall
by the ookinetes of Plasmodium gallinaceum. J
Protozool 39: 449-454.
Torii M, Yanagi T, Tsuboi T, Kanbara H 1992b. Appearence and
localization of Plasmodium yoelii circumsporozoite
protein during sporogony in Anopheles stephensi. J
Protozool Res 2: 134-140.
Touray MG, Warburg A, Laughinghouse A, Krettli AU, Miller LH
1992. Developmentally regulated infectivity of malaria
sporozoites for mosquito salivary glands and the vertebrate
host. J Exp Med 175: 1607-1612.
Tsuji M, Corradin G, Zavala F 1992. Monoclonal antibodies
recognize a processing dependent epitope present in the mature
CS protein of various plasmodial species. Parasite
Immunol 14: 457-469.
Turner DP, Gregson NA 1982. The cell surface of Plasmodium
gallinaceum sporozoites: microelectrophoretic and lectin-
binding studies. Parasitology 84: 227-238.
Van Houten J 1988. Chemoresponse mechanism toward a molecular
level. J Protozool 35: 241-245.
Vanderberg JP 1974. Studies on the motility of
Plasmodium sporozoites. J Protozool 21:
527-537.
Vanderberg JP 1975. Development of infectivity of the
Plasmodium berghei sporozoite. J Parasitol
61: 43-50.
Vanderberg JP 1977. Plasmodium berghei: quantitation of
sporozoites injected by mosquitoes feeding on a rodent host.
Exp Parasitol 42: 169-181.
Vanderberg JP, Rhodin J, Yoeli M 1967. Electron microscopic
and histochemical studies of sporozoite formation in
Plasmodium berghei. J Protozool 14: 82-
103.
Vaughan JA, Do Rosario V, Leland P, Light J, Woolett R,
Hollingdale MR, Azad AF 1988. Plasmodium falciparum:
ingested anti-sporozoite antibodies affect sporogony in
Anopheles stephensi mosquitoes. Exp Parasitol
66: 171-182.
Vaughan JA, Noden BH, Beier JC 1992. Population dynamics of
Plasmodium falciparum sporogony in laboratory-infected
Anopheles gambiae. J Parasitol 78: 716-724.
Vaughan JA, Noden BH, Beier JC 1994. Sporogonic development of
cultured Plasmodium falciparum in six species of
laboratory-reared Anopheles mosquitoes. Am J Trop
Med Hyg 51: 233-243.
Vermeulen AN, Deursen JV, Brakenhof RH, Lensen THW, Ponnudurai
T, Meuwissen JHET 1986. Characterization of Plasmodium
falciparum sexual stage antigens and their biosynthesis in
synchronized gametocyte cultures. Mol Biochem
Parasitol 20: 155-163.
Vermeulen AN, Ponnudurai T, Beckers PJA, Verhave JP, Smits MA,
Meuwissen JHET 1985a. Sequential expression of antigens on
sexual stages of Plasmodium falciparum accessible to
transmission blocking antibodies in the mosquito. J Exp
Med 162: 1460-1476.
Vermeulen AN, Roeffen WFG, Hendrik JBJ, Ponnudurai T, Beckers
PJA, Meuwissen JHET 1985b. Plasmodium falciparum
transmission blocking antibodies recognize monovalently
expressed epitopes. Develop Biol Standard 62: 91-97.
Vernick KD, Collins H, Gwadz RW 1989a. A general system of
resistance to malaria infection in Anopheles gambiae
controlled by two main genetic loci. Am J Trop Med Hyg
40: 585-592.
Vernick KD, Collins FH 1989b. Association of a
Plasmodium-refractory phenotype with an esterase locus
in Anopheles gambiae. Am J Trop Med Hyg
40: 593-597.
Vickerman K, Cox FEG 1972. Introductory studies in Biology. In
SW Hurry & J Murray (eds), The Protozoa, Albemarie
Street, London.
Warburg A, Miller LH 1992. Sporogonic development of a malaria
parasite in vitro. Science 255: 448-
450.
Warburg A, Schneider I 1993. In vitro culture of the mosquito
stages of Plasmodium falciparum. Exp Parasitol
76: 121-126.
Warburg A, Touray M, Krettli AU, Miller LH 1992. Plasmodium
gallinaceum: antibodies to circumsporozoite protein
prevent sporozoites from invading the salivary glands of
Aedes aegypti. Exp Parasitol 75: 303-
307.
Weathersby AB 1952. The role of the stomach wall in the
exogenous development of Plasmodium gallinaceum as
studied by means of haemocoel injections of susceptible and
refractory mosquitoes. J Infec Dis 91: 198-
205.
Weathersby AB 1954. The ectopic development of malarial
oocysts. Exp Parasitol 3: 538-543.
Weathersby AB 1960. Experimental infection of Aedes
aegypti with exoerythrocytic stages of Plasmodium
gallinaceum. Exp Parasitol 9: 334-337.
Wernsdorfer WH 1991. The development and spread of drug-
resistant malaria. Parasitol Today 7: 297-
303.
Wilkinson RN, Colwell EJ, Neoypatimanond S 1973. Effect of
sulphamethoxazole-trimethoprim on the viability of
Plasmodium falciparum gametocytes. Trans R Soc Trop
Med Hyg 67: 148-149.
Wilkinson, RN, Neypatimanond, S, Gould DJ 1976. Infectivity of
falciparum malaria patients for anopheline mosquitoes befoore
and after chloroquine treatment. Trans R Soc Trop Med
Hyg 70: 306-307.
Winger LA, Tirawanchai N, Nicholas J, Carter HE, Smith JE,
Sinden RE 1988. Ookinete antigens of Plasmodium
berghei. Appearance on the zygote of a Mr 21 kD
determinant identified by transmission blocking monoclonal
antibodies. Parasite Immunol 10: 193-207.
Wirtz RA, Burkot TR, Graves PM, Andre RG 1987. Field
evaluation of enzyme-linked immunosorbent assays for
Plasmodium falciparum and Plasmodium vivax
sporozoites in mosquitoes (Diptera: Culicidae) from Papua New
Guinea. J Med Entomol 24: 433-437.
Yoshida N, Potonjak P, Nussenzweig V, Nussenzweig RS 1981.
Biosynthesis of Pb44, the protective antigen of sporozoites of
Plasmodium berghei. J Exp Med 154:
1225-1236.
Young MD, Burgess RW 1961. The infectivity to mosquitoes of
Plasmodium malariae. Am J Hyg 73: 182-
192.
The Biology of Malarial Parasite in the Mosquito - A
Review
Amauri Braga Simonetti
Departamento de Microbiologia, Instituto de Biociencias,
Universidade Federal do Rio Grande do Sul, Rua Sarmento Leite
500, 90050-170 Porto Alegre, RS, Brasil
Received 20 December 1995
[TABLES AND FIGURES AT END OF TEXT]
The purpose of this review is to summarize the biology of
Plasmodium in the mosquito including recent data to
contribute to better understanding of the developmental
interaction between mosquito and malarial parasite. The entire
sporogonic cycle is discussed taking into consideration
different parasite/vector interactions and factors affecting
parasite development to the mosquito.
Key words: malaria - mosquito - sporogonic cycle - cell
biology
The control and prevention of malaria has been pursued for a
long time. Although campaigns against malaria were initially
successful in some areas, the emergence of resistance of the
parasite to drugs and of the mosquito vector to insecticides,
combined with the difficulties in implementing and maintaining
effective control schemes have led to a resurgence of the
disease in many parts of the world (Wernsdorfer 1991, Schapira
et al. 1993, Roush 1993). Much of the current work on malaria
focuses on immunological aspects of the disease and there has
been remarkable progress in the identification of a variety of
parasite antigens in different stages of the parasite
development, some of which may be included in a future vaccine
(Nussenzweig & Nussenzweig 1989, Mitchel 1989, Targett 1989,
Nussenzweig 1990, Hommel 1991, Good 1991a). However,
experimental and field studies have shown the complexity of
the immune response to parasites, indicating that an efficient
malaria vaccine is difficult to achieve (Cattani 1989, Good
1991b, 1992, Philips 1992). Therefore, for control of the
disease greater understanding of the biological mechanisms
involved in host/parasite/vector interactions is essential.
In recent years an increasing number of studies have
concentrated on the mosquito stages of the parasite
development. Comprehensive reviews on biological,
ultrastructural, biochemical, molecular and immunological
aspects of the parasite and the vector can be obtained in the
literature (Sinden 1978, 1983, 1984, Carter & Graves 1988,
Carter et al. 1988, Alano & Carter 1990, Billingsley 1990,
Crampton et al. 1990, 1994, Alano 1991, Brey 1991, Coluzzi
1992, Kaslow et al. 1994a, Sinden et al. 1996). This review
foccuses on the cell biology of malarial parasites at
different stages of its development in the mosquito and also
includes factors that may affect the parasite infectivity.
The sporogonic cycle
Some erythrocytic parasites differentiate into sexual forms
called gametocytes. Mature and functional gametocytes ingested
by an appropriate species of mosquito in a bloodmeal are
stimulated to transform into the stages which establish the
parasites in their vector (Garnham 1966). Under the influence
of changes in the mosquito midgut environment the gametocytes
become extracellular within 8-15 min of ingestion. After
emergence from the red blood cell (exflagellation) the male
gametes fertilize the female gametes within 60 min of
ingestion of blood. The fertilized macrogamete (zygote)
differentiates into a single motile ookinete over the next 10-
25 hr, which migrates from the bloodmeal through the midgut
wall to form an oocyst underneath the basal lamina of the
midgut. Each oocyst produces many thousands of invasive
sporozoites over a period of 7-12 days. The sporozoites escape
from the oocyst and then invade the salivary glands, here they
stay for possibly very long periods until injected into
another vertebrate host when the next bloodmeal is taken
(Sinden 1984, Carter & Graves 1988). A diagram of the
sporogonic cycle in the mosquito is shown in Figure.
Development of gametocytes (gametocytogenesis)
As with other members of the order Haemosporina, gametocytes
of Plasmodium can arise from merozoites from pre-
erythrocytic parasites. Nevertheless, during natural
infections, most gametocytes arise from merozoites of blood-
stage origin (Alano & Carter 1990). Two alternative
possibilities by which malaria parasites could become
committed to either sexual or asexual development have been
proposed (Inselburg 1983, Mons 1985, 1986, Bruce et al. 1990):
first, the merozoite is not committed at the time of invasion
of a red blood cell. During early growth (as a ring form), the
parasite is suceptible to factors that will commit it to
either sexual or asexual development. Second, during growth of
an asexual parasite, environmental factors influence it so
that at maturity the schizont produces merozoites that are
precommitted to form asexual parasites or committed to form
gametocytes upon subsequent invasion of a red blood cell.
A number of studies have attempted to identify factors that
may influence the differentiation and production of sexual
stage parasites including cAMP, presence of antimalarial drugs
and other environmental factors associated to the host such as
the presence of serum undetermined components (reviewed by
Sinden 1983, Mons 1986, Carter & Graves 1988, Dearsly et al.
1990). In a recent report (Schneweis et al. 1991) it was
claimed that the production of infective gametocytes in
vitro can be enhanced by products of haemolysis but the
nature of the erythrocytic factor was not identified.
Gametocyte development takes longer than asexual schizogony,
e.g. 26 hr as opposed to 22.5 hr in P. berghei (Mons et
al. 1985) or in the more extreme case of P. falciparum
8-12 days as opposed to 48 hr (Sinden 1983). The proportion
of parasites that develop into gametocytes varies greatly
during the course of natural infections even at its peak but
is very low in relation to the total parasitaemia (Smalley et
al. 1981). The growth and differentiation of gametocytes of
P. falciparum has been divided into five stages (I to
V) covering about eight days from merozoite invasion to mature
gametocyte, each stage being distinguished by successive
changes in the organization of the cell (Hawking et al. 1971).
Little is known about commitment of gametocytes to be male or
female. The fact that both female and male can be produced by
haploid blood-stage parasite, the sex of a gametocyte does not
result from chromosomal differences between both types of cell
but their development must be due to selective gene
expression. In general, the number of female gametocytes
predominates over the number of male, but this predominance
may vary at different times between cloned infections
(Cornelissen 1988, Schall 1989).
Gametogenesis
Mature and functional gametocytes ingested by the mosquito in
a bloodmeal are stimulated by the midgut environment to
transform into gametes. Studies indicate that various triggers
induce gametocytes to undergo differentiation. Microgameto-
genesis in vitro is, optimally, dependent upon a rise
in pH (Nijhout & Carter 1978), a rise in pCO2 and bicarbonate
levels (Carter & Nijhout 1977, Nijhout & Carter 1978), a fall
in temperature of a few degrees below that of the vertebrate
host (Sinden & Croll, 1975) or a very potent factor termed
mosquito exflagellation factor (MEF). The latter is a small
heat stable molecule from the mosquito head and gut which
stimulates gametogenesis via a bicarbonate- and pH-independent
mechanism (Nijhout 1979). Recently, Kawamoto et al. (1991)
showed in vitro that induction of exflagellation of
P. berghei is triggered by a rise in the intracellular
pH (pHi) which is mediated by Ca^++ and cGMP regulation. pHi
can be modulated by alkaline media and is controlled by a
complex series of interdependent ion pumps and channels
controlling Na^+, K^+, Cl^- and HCO3^- transport between the
parasite and the environment. Other influential factors
described include cAMP analogues and inhibitors of
phosphodiesterase (Martin et al. 1978). The duration of
microgametogenesis is both temperature and species dependent,
e.g. at 20-22 C it may take 7-15 min for P. falciparum
in vitro, although exflagellation may be detected after
shorter periods in the fluid excreted by feeding
Anopheles (Sinden 1983). There is no evidence that
exflagellation is influenced by factors released by digestion
of the blood meal since digestion normally begins several
hours later (Graf et al. 1986). Microgamete formation involves
three mitotic divisions with a rapid assembly of eight
axonemes on the single microtubule organizing centre that
divides and passes to the spindle poles. This division
simultaneously segregates the genome and the axoneme so that
each of the eight emergent gametes receives a single axoneme
and haploid genome, both being connected to a common
microtubule organizing centre. After exflagellation the
microgametes, normally bearing a single axoneme, a single
condensed nucleus and a single kinetosome with its sphere and
granule at the distal end, are torn from the microgametocyte
surface and rapidly move away into the blood meal (Sinden &
Croll 1975).
Macrogametogenesis at the morphological level involves little
more than escape from the host cell (Sinden 1984). At the
cellular level there is de novo synthesis of the
proteins which are expressed on the surface of macrogamete
(Kumar & Carter 1984). It was recently identified a gametocyte
specific protein of P. falciparum called Pf11-1 and
there is some evidence that this protein might be involved in
the emergence of gametes from the infected erythrocyte (Scherf
et al. 1992, 1993).
Interactions between the vertebrate host and the parasite do
not cease when the blood meal is taken. Following induction of
gametogenesis the parasite is liberated into the blood meal.
It has been shown that the gametes are susceptible to the
phagocytes in the host's blood (Rutledge et al. 1969, Sinden &
Smaley 1976) and that they are susceptible to activation of
the components of the complement pathway (Grotendorst et al.
1986, 1987).
Zygote formation (fertilization) and ookinete
development
Fertilization is rapid usually occurring within 1 hr of gamete
formation, the plasmalemma of the two gametes fuse and the
microgamete axoneme and nucleus enter the macrogamete
cytoplasm (Sinden 1983). During zygote development, structural
changes occur and its transformation to ookinete is in part
determined by the gradual assembly of the apical complex
including the collar and pollar rings, rhoptries and
micronemes (Canning & Sinden 1973, Davies 1974, Sinden 1978).
Intensive protein synthesis also begins in the fertilized
macrogamete and continues in the zygote and developing
ookinete as reported for different Plasmodium species
(Kaushal et al. 1983a, Kaushal & Carter 1984, Kumar & Carter
1985, Vermeulen et al. 1985a, 1986, Sinden et al. 1987).
Included in this repertoir of proteins are many of the targets
of proposed transmission blocking immunity. It has been
reported that An. gambiae can concentrate the bloodmeal
by a factor of 1.2 to 1.8 reducing the production of ookinetes
when compared to An. freeborni which can not
concentrate (Sinden et al. 1996). The mature ookinete is a
motile cell that varies from 7 to 18 mm in length and 2 to 4
mm in diameter (Sinden, 1978). Locomotion of the ookinete has
been described in different Plasmodium species as a
linear or snake-like gliding motion (Freyvogel 1966). Studying
P. berghei ookinetes in primary mosquito cell cultures,
Speer et al. (1974) described spiral waves on the surface of
some ookinetes, especially in the anterior half of the body,
which might be involved in ookinete locomotion.
Some factors could influence the development, survival and
infectivity of the parasite during its residence in the midgut
lumen. Eyles (1952) has showed that the parasite development
ceases at the ookinete stage unless a macromolecular (non-
dialyzable) component is present in the blood meal. Studying
the influence of red blood cells on the ability of P.
gallinaceum zygotes fertilized in vitro to infect
Aedes aegypti Rosenberg et al. (1984) found a linear
relationship between erythrocyte density and the number of
oocysts up to a 50% hematocrit. Furthermore, they deduced that
there are one or more nondialyzable substances (erythrocytic
factors) contained in normal erythrocytes, and released by
mosquito digestion, that are essential for ookinete invasion
of the gut epithelium. When they added trypsin inhibitor to
the bloodmeal there was an inhibition of midgut penetration by
ookinetes. Recent studies have shown that when mosquitoes are
fed with cultured P. falciparum (Ponnudurai et al.
1989) and P. berghei (Sinden 1989) gametocytes, upon
dilution with fresh red cells, more oocysts result at initial
(low) dilutions whereas further dilution reduces oocyst
counts. The involvement of blood factors and/or its digestive
products in infectivity has been studied in different
parasite-vector models. Using a selected line of An.
stephensi, Feldmann and Ponnudurai (1989) found mature
P. falciparum ookinetes in the midgut lumen of refractory
mosquitoes but no further penetration of the gut epithelium
was observed. The reasons for this limited development in non-
compatible mosquitoes could be related to digestive function
since early initiation of hemoglobin degradation and higher
aminopeptidase activity have been described in refractory
strains of An. stephensi (Feldmann et al. 1990). It has
also been shown that P. gallinaceum develops up to the
ookinete stage in the non-compatible mosquito An.
stephensi, this development occurring over the same time
period and with the same success as in the compatible vector
Ae. aegypti. However, P. gallinaceum ookinetes
did not escape from the midgut lumen in An. stephensi
mosquitoes (Rudin et al. 1991).
Possible mechanism inhibiting parasite development involves
damage of the parasite by digestive enzymes present in the
vector. Trypsin and aminopeptidases are the major proteolytic
enzymes involved in blood digestion by female mosquitoes, and
are produced by the midgut cells in direct response to blood
feeding (Briegel & Lea 1975, Graf & Briegel 1982, Billingsley
1990, Billingsley & Hecker 1991). P. gallinaceum
ookinetes 0-10 hr old (i.e. zygote to ookinete transition)
were shown to be susceptible to mosquito enzymes in double
feeding experiments (Gass 1977) and in vitro damage was
observed to cultured ookinetes by proteinases from Ae.
aegypti (Gass & Yeates, 1979). However, results found by
Shahabuddin et al. (1993) using the same parasite/vector
system suggest that the parasite secretes an inactive or
partially active chitinase that is activated by a mosquito-
produced serine protease. In a recent study Chege et al.
(1996) examined the effect of digestive enzymes on the
kinetics of P. falciparum ookinete development and
oocyst infection rates in An. albimanus, An.
freeborni and An. gambiae. Their data indicated
that proteolytic enzymes alone do not limit the early stages
of sporogonic development in these vector species of
Anopheles.
Peritrophic layer
The peritrophic layer (PL) of the insect midgut forms a
cylindrical sheet separating the midgut contents from the
single cell-layered midgut endothelium. It is secreted within
hours of the blood meal at different rates depending on the
mosquito species (Billingsley 1990). Apical secretion granules
are present in the midgut cells of Anopheles species,
which are released into the periphery of the posterior midgut
lumen during the feeding process in response to stretching of
the gut wall. The vesicle contents coalesce and then condense
to form a compacted PL between 8 and 24 hr (Hecker 1977,
Berner et al. 1983). In culicine mosquitoes the PL is formed
de novo by the posterior midgut cells in the
proventriculus. In A. aegypti, formation starts
immediately after the blood meal, but about 12 hr later it is
a mature structure (Perrone & Spielman, 1988).
The role of the mosquito PL as a barrier to ookinete invasion
of the gut wall and pathogenic effects of Plasmodium
species upon the vector are controversial. In ultrastructural
studies on the interaction of P. falciparum ookinetes
with the midgut epithelium of An. stephensi Meis &
Ponnudurai (1987) frequentely found parasites trapped in the
membrane. They also observed that if the PL was dissected out
36 hr after the blood meal many ookinetes were attached to its
external surface. In the same study it was mentioned that the
ookinete was capable of penetrating the newly formed, but not
the thickened and hardened PL 36 hr after the feeding. A
failure to cross this barrier or retarded penetration might
increase the length of exposure of ookinetes to mosquito
trypsin to which it is known to be sensitive. In contrast,
using P. berghei-infected An. atroparvus
mosquitoes, Sluiters et al. (1986) concluded that the PL would
not function as physical barrier against migrating ookinetes
which can pass through fenestrations. Billingsley and Rudin
(1992) observed that infectivity of An. stephensi by
P. berghei, measured by oocyst counts, was unnaffected
by the presence or absence of the PL. However, in A.
aegypti infected with P. gallinaceum its presence
serves to reduce rather than prevent infection (Ponnudurai et
al. 1988, Billingsley & Rudin 1992). These observations
suggested to the authors that in compatible vector-parasite
combinations, the PL acts as a limiting, rather than absolute
barrier to the penetrating ookinete while in incompatible
combinations the PL appears to be an absolute barrier to
ookinete penetration.
To reach the midgut epithelium the ookinetes must first cross
the partially or fully formed PL. The PL may act as the
recognition site for the penetrating ookinete via lectin-
mediated mechanisms. The occurrence of sugar residues which
could be involved in vector recognition by the parasite has
been demonstrated in the PL of An. stephensi and Ae.
aegypti (Berner et al. 1983). Rudin and Hecker (1989)
showed the presence of binding sites for different lectins in
midguts of P. berghei-infected An. stephensi,
with high specificity for N-acetyl-D-galactosamine
(GalNAc), and P. gallinaceum-infected Ae. aegypti,
with high specificity for N-acetyl-D-glucosamine (GlcNAc).
The authors concluded that it seems likely that lectin-binding
phenomena play a role in the orientation of the parasites on
their way out of the midgut lumen and that the PM and/or
glycocalyx may be crucial structures for the penetration of
the gut epithelium by the ookinete. This does not exclude the
possibility that ookinetes penetrate the PL by an enzymatic
process (see below). Ultrastructural observations on Ae.
aegypti infected with P. gallinaceum showing an
electron dense amorphous material in front of the parasite
were consistent with a blockade of parasites within the PL
(Sieber et al. 1991). The ookinetes appeared to disrupt the
layers of the PL, suggesting an enzymatic mechanism for
penetration. These observations were further investigated by
Huber et al. (1991) who, using the same vector/parasite
system, also suggested a possible role for GlcNAc in the
binding of the ookinete to the PL, and demonstrated the
presence of chitin in the PL. They observed that mature
ookinetes transiently secreted a soluble chitinase, thought to
be responsible for the digestion of the PL. Recently,
Shahabuddin et al. (1993) reported an inhibition of P.
gallinaceum chitinase and a transmission blocking activity
of a chitinase inhibitor, allosamidin, on the sporogonic
development of P. gallinaceum and P. falciparum
in the respective vectors Ae. aegypti and An.
freeborni. However, enzymatic mechanisms of penetration
may differ in other vector/parasite systems since the PL of
An. stephensi appears not to contain chitin (Berner et
al. 1983).
Penetration of midgut epithelium
Ookinetes found in the epithelial cell layer between 24 and 48
hr post-infection have successfully evaded the obstacles
presented by the midgut lumen; however, they still might fail
to develop at the normal site of development (Sinden 1984,
Ponnudurai et al. 1988). It has been shown that the midgut
wall is negatively charged (Houk et al. 1986), but there is no
evidence of electrical charge interactions between ookinetes
and epithelial cells. It has been a matter for discussion as
to whether the ookinete follows an intra- or intercellular
route to reach the ultimate site of development and encystment
in the outer wall of the midgut epithelium.
Intercellular movement of P. gallinaceum ookinetes was
first described by Stohler (1957), but Mehlhorn et al. (1980)
have found the same parasite in an intracellular position.
Recently, Torii et al. (1992a) observed P. gallinaceum
ookinetes in both intracellular and intercellular
positions in the midgut epithelium of the mosquito Ae.
aegypti, which they interpret as that they first enter
into the epithelium, then exit into the intercellular space
and move to the basal lamina. Garnham et al. (1962) showed
that the ookinete of P. cynomolgi bastianelli enters
the epithelial cell by liquifying the cell membrane. Davies
(1974) again postulated intercellular movement by P.
berghei nigeriensis ookinetes. Although describing the
ookinete of P. berghei in an intracellular position, as
did Garnham et al. (1969), Canning and Sinden (1973) stated
that the parasite might also migrate by an intercellular
route. More recently, P. yoelii nigeriensis ookinetes
have been described to take an intracellular route to the
external wall of the midgut (Maier et al. 1987). However, when
the same parasite was used to infect An. omorii, an
intercellular route was mostly undertaken, although the
intracellular occurrence was also observed (Syaffruddin et al.
1991). Meis and Ponnudurai (1987) presented evidence that
P. falciparum ookinetes migrate between the epithelial
midgut cells. Using a specific monoclonal antibody they also
observed a track in the PL, which is related to the shedding
of a 25 kD surface protein (Pfs25) during movement. The
authors suggested that this protein may bind to receptors on
the epithelial cells prior to an intercellular invasion, since
it is reported to have epidermal growth factor (EGF)-like
domains (Kaslow et al. 1988). It was recently shown that Pfs25
persits in the oocyst wall during parasite development in the
mosquito (Lensen et al. 1993). The same group studied the
migration of P. falciparum and P. berghei
ookinetes through the midgut epithelium in An.
stephensi by using ruthenium red staining (Meis et al.
1989). The results of previous studies were confirmed: P.
falciparum ookinetes penetrated by intercellular route,
but the rodent parasite P. berghei appeared to take an
intracellular position, confirming that both mechanisms occur
and are species-dependent. In the case of P. berghei a
protein of 21 kD (Pbs21) present on the surface of the
ookinete (Sinden et al. 1987) could play a role during the
intracellular invasion of the midgut epithelium of An.
stephensi mosquitoes. It was demonstrated in the midgut of
P. berghei infected mosquitoes that expression of Pbs21
was predominantly localized on the ookinete surface one day
after the infectious blood meal and thereafter expression
declined to a minimum on days 2 and 3, the time of onset of
oocyst development (Simonetti et al. 1993).
The mode of penetration by ookinetes can perhaps be related to
damage of the epithelial lining of the midgut. Intercellular
migration may not damage cell membranes, and increased
mortality does not occur in P. falciparum-infected
An. stephensi and An. gambiae during this
period, even with very heavy parasite loads (Meis &
Ponnudurai, 1987). Similar observations have been reported in
P. gallinaceum-infected Ae. aegypti (Freier &
Friedman, 1987). They observed similar mortality rates in
infected and uninfected batches of mosquitoes. However, when
ookinetes use an intracellular route, as described previously,
increased damage to midgut cells might occur, resulting in
higher mosquito mortality. This is probably mediated by
invasion of the hemolymph by opportunistic gram-negative
bacteria and/or microsporidia (Maier et al. 1987, Seitz et al.
1987).
Ultimately the ookinete penetrates the basement membrane, but
fails to pass through the basal lamina of the midgut adjacent
to hemocoele (Sinden 1984). Whether this is due to the
inability of the ookinete to penetrate the basal lamina, or to
the specific recognition of the lamina and consequent shut-
down of the incisive process is not known. Interactions of
parasites with vector extracellular matrix proteins (ECM)
cannot be discounted (Kaslow et al. 1994b).
Oocyst development
Oocyst development is predominantly extracellular (Duncan et
al. 1960, Garnham et al. 1969, Howells & Davies 1971, Sinden
1975), but occasionally occurs within the midgut epithelial
cell (Vanderberg et al. 1967, Bafort 1971, Beaudoin et al.
1974). The ookinete usually comes to rest beneath the basal
lamina 18-24 hr after the infective blood meal (Sinden 1978).
It rapidly rounds up between 18 and 72 hr after feeding and
the apical complex is resorbed into the oocyst cytoplasm
(Garnham et al. 1969). There is some evidence suggesting a
significant role of the basal lamina in the development of the
ookinete (Kaslow et al. 1994b). It was found that in vitro-
cultured ookinetes injected directly into the hemolymph
form clusters of oocysts adherent to the basal lamina
throughout the hemocoele. Furthermore, binding of ookinetes to
artificial surfaces, such as plastic, is enhanced at least 10-
fold by addition of various components of basal lamina such as
matrigel, collagen IV, and laminin (Warburg & Miller, 1992).
The young oocyst is enveloped by a thick plasmalemma that is
covered on the hemocoelomic surface by a fibrous basal lamina.
Oocysts from the second day onwards are also covered by an
amorphous capsule which becomes reduced in thickness at
maturity (Vanderberg et al. 1967, Aikawa 1971, Strome &
Beaudoin 1974, Sinden 1975).
Despite the usual growth of the oocyst under the basal lamina
of the midgut, oocyst development is not site specific.
Weathersby (1952, 1954, 1960) has shown by injection of
gametocytes directly into the hemocoele of susceptible
mosquitoes that oocysts would develop to maturity if attached
to other parts of the body than the stomach or even if they
were floating freely in the hemocoel fluid. In his experiments
he used different parasite-host combinations and concluded
that the site of oocyst development is probably not a critical
factor in the maturation process. Furthermore, the factors
that are responsible for the death of parasite in refractory
lines are not confined to the stomach wall. These results were
supported by those reported by Ball and Chao (1957, 1960,
1961, 1976) who showed that oocysts of P. relictum may
develop in vitro away from the intact stomach of the
mosquito. The overall results of this series of studies by
Ball and Chao demonstrated in vitro development of all
stages from ookinetes to fully infective sporozoites without
attachment of oocysts to the midgut. However, it was not
possible to obtain complete sporogonic development in a single
preparation. Rosenberg and Koontz (1984) injected cultured
P. gallinaceum zygotes into the hemocoele of Ae.
aegypti mosquitoes and observed development of ectopic
oocysts in approximately 50% of the mosquitoes, with
sporozoites being found in the salivary glands. These
observations suggest that oocyst metabolism is not dependent
upon direct transfer of nutrients from the midgut epithelium.
Occasional intracellular oocyst development has been reported
for P. berghei in An. stephensi and An.
quadrimaculatus (Vanderberg et al. 1967). The same
localization was described in P. vinckei by Bafort
(1971) who concluded that both mechanical pressure and
physiological mechanisms play a role in the movement of
oocysts to the hemocoelomic surface. Studying the sporogonic
development of P. berghei in An. stephensi,
Beaudoin et al. (1974) found oocysts developing ectopically
within the midgut epithelium following normal infection,
eventually emptying their sporozoite content into the tissue
itself or the midgut lumen. In addition, they observed no
morphological and structural abnormalities in the luminal
parasites which displayed good viability. In contrast to P.
berghei, no ectopic development was seen in P.
falciparum-infected An. stephensi mosquitoes (Meis
et al. 1992b), confirming previous results observed with P.
falciparum in naturally infected An. gambiae
(Sinden & Strong 1978). Recent reports described an
enhancement of oocyst development in vitro for P.
berghei (Syaffruddin et al. 1992), P. gallinaceum
(Warburg & Miller, 1992) and P. falciparum (Warburg &
Schneider, 1993), when insect cell lines were added into the
culture medium. From these observations it appears that
nutritional or other regulatory requirements of the developing
parasite can be met without a direct contact with midgut
epithelium or haemolymph. The question of how the oocyst is
supplied with nutritive material is an intriguing one.
Little information is available on the uptake and source of
nutrients for oocyst development. It is assumed that in
vivo the source of nutrients is the hemolymph. Mack and
Vanderberg (1978) analyzed hemolymph of An. stephensi
collected from uninfected and P. berghei-infected
mosquitoes at different stages of the parasite development. It
was found that four days after the blood meal the osmotic
pressure and the specific gravity were lower in infected
mosquitoes compared with uninfected ones. The difference,
however, was attributed to indirect effects of the quality of
the ingested blood meal. These studies were complemented by
analysis of the concentration of free amino acids in the
hemolymph collected in similar conditions with results showing
significantly lower concentrations in infected mosquitoes with
decreases in valine and histidine, and a total loss of
detectable methionine suggesting it is incorporated (Mack et
al. 1979). This difference could be due to the utilization of
some of these amino acids by the developing oocyst as
suggested by Ball and Chao (1976) who analyzed the uptake of
amino acids by P. relictum oocysts in vitro,
comparing growth of uninfected and infected guts of Culex
tarsalis in Grace's insect culture medium. They found
significant decreases in the concentration of certain amino
acids including arginine, asparagine, proline and histidine,
and less marked decrease in concentrations of others like
methionine, valine, leucine and isoleucine. From these studies
it appears that the reduced amount of free aminoacids in the
hemolymph is due to oocyst metabolism. Autoradiographic
studies with P. gallinaceum in Ae. aegypti
mosquitoes indicated that ^3H-leucine is uniformly
incorporated throught the oocyst within 15 min of injection
into hemocoele (Vanderberg et al. 1967).
Sporogony
With increasing maturation the oocyst undergoes considerable
cytoplasmic subdivision. Initially the plasmalemma forms
invaginations and clefts that penetrate even deeper into the
cytoplasm, thus subdividing the cell (Vanderberg et al. 1967,
Terzakis 1971, Posthuma et al. 1988). In a transmission
electron microscopy study of P. falciparum oocysts it
was suggested that cleft formation was due to dilation of
endoplasmic reticulum (Sinden & Strong 1978). Using immunogold
labelling technique during sporogonic stage of the same
parasite, Posthuma et al. (1988) considered the latter
explanation unlikely. With increasing activity the cytoplasmic
clefts become extended and the expanding vacuolar space more
pronounced leading to the sporoblast formation. Along the
clefts sporozoites are formed by a budding process at the
surface of the limiting membrane (Vanderberg et al. 1967,
Howells & Davies 1971, Canning & Sinden 1973, Sinden & Strong
1978).
As the sporozoite continues to bud off, a nucleus and various
cytoplasmic components are passed into it from the sporoblast.
The membranes of the developing sporozoite pellicle are formed
and other organelles like microtubules and rhoptries become
discernible (Vanderberg et al. 1967, Sinden & Garnham 1973).
When sporogony is completed (about 10-12 days after the
infective feed), the oocyst is filled with sporozoites and one
or more residual bodies (Sinden 1984). Estimations of number
of sporozoites per oocyst have ranged widely. Garnham (1966)
reported that the number in a single P. vivax oocyst
varies from 1,000 to 10,000. Pringle (1965) estimated that a
single oocyst of P. falciparum contains nearly 10,000
sporozoites. Studies carried out with mosquitoes fed on
infected volunteers from Thailand showed a mean count of
approximately 3,700 sporozoites per oocyst for P.
vivax and 3,400 for P. falciparum, whereas for
P. cynomolgi-infected mosquitoes a single oocyst
contained about 7,500 (Rosenberg & Rungsiwongse 1991).
Dependent upon species, the mature sporozoite varies from 9 to
16.5 mm in length and from 0.4 to 2.7 mm in diameter; aberrant
forms have been described up to 40 mm long (Sinden 1978).
So far, studies on synthesis and expression of proteins during
sporogonic development have foccused on a polypeptide called
the circumsporozoite protein (CSP) found on the mature
sporozoite. Observations on the origin of CSP and its
distribution through the mosquito stage were reported by
several authors. It is now well established that these
proteins are synthesized in maturing oocysts of different
Plasmodium species from 6-7 days after the infective
blood meal, before sporozoites are visible (Nagasawa et al.
1987, 1988, Posthuma et al. 1988, Hamilton et al. 1988,
Boulanger et al. 1988, Torii et al. 1992b, Meis et al. 1992a).
At this stage, CSP is present on the plasmalemma and at
various sites within the cytoplasm and endoplasmic reticulum
of the sporoblast. When the sporozoites bud from the
sporoblast they are already covered with CSP which is also
found in salivary gland sporozoites (Yoshida et al. 1981,
Santoro et al. 1983, Tsuji et al. 1992).
Humoral encapsulation of oocysts, which in malaria infected
mosquitoes is known as Ross' black spores, has been described
in P. berghei nigeriensis and P. vivax and seems
to occur mainly in older oocysts which have begun to produce
sporozoites (Sinden & Garnham, 1973). This phenomenon was
studied in a selected line of An. gambiae that
encapsulates different Plasmodium species (Collins et
al. 1986). The authors demonstrated that refractoriness is
manifested by melanization of the ookinete after its passage
through the midgut epithelium. Paskewitz et al. (1989)
localized phenoloxidase activity in the basal lamina of the
epithelial cells of both encapsulating and susceptible
mosquitoes prior to blood feeding. However, after an infective
blood meal, this activity was still observed close to invading
ookinetes in refractory mosquitos but it was reduced or absent
in susceptible mosquitoes. When the non-compatible vector
An. gambiae was fed with ookinetes of P.
gallinaceum, invasion of the midgut epithelium by the
ookinetes occurred but oocysts were infrequentely formed.
Using the same system Vernick et al. (1989) and Vernick and
Collins (1989) tried to elucidate mechanisms involved in
vector-parasite incompatibility by injecting in vitro-
cultured ookinetes into the hemocoel of mosquitoes and
monitoring parasite development using specific rRNA probes. As
no differences were found between susceptible and refractory
mosquitoes the authors suggested that the specific lytic
factor(s) in the refractory line are intracellular.
The sporozoite and salivary gland invasion
The motile sporozoites emerge into the hemocoele through holes
from an area of weakness in the oocyst wall. Holes are
possibly produced by a combination of the muscular action of
the gut wall and the activity of the sporozoites (Sinden 1978,
Meis et al. 1992b). Within the hemocoel the sporozoites are
distributed throughout the mosquito and can initially be found
in many parts of the insect, even in the maxillary palps;
within a day or two of their release from oocysts the
sporozoites invade the salivary glands where they accumulate
and remain until delivery (Vaughan et al. 1992). Thus,
sporozoites do not adhere to most tissues, except for the
salivary glands and rarely the midgut wall, hemocytes or
thoracic muscles (Vanderberg 1974, Sinden 1975, 1978, Golenda
et al. 1990, Vaughan et al. 1992). The latter observation,
especially in heavily infected mosquitoes, could be associated
with an impairment of flight activity in malaria-infected
An. stephensi vectors demonstrated by some authors
(Rowland & Boersma 1988).
It has been estimated that in mosquitoes fed on individuals
with naturally acquired P. vivax about 850 sporozoites
per oocyst reached the glands (Sattabongkot et al. 1991) which
follows, by calculation, that approximately 23 % of all P.
vivax sporozoites released into the hemocoel subsequentely
reach the salivary glands (Rosenberg & Rungsiwongse 1991).
Vaughan et al. (1992), using regression analysis, calculated
the approximately 650 salivary gland sporozoites were produced
per oocyst and reported that virtually all oocyst infections
produced salivary gland infections in An. gambiae
infected with P. falciparum by membrane feeding. The
same group has found a similar number by studying the
sporogonic development of cultured P. falciparum in six
species of laboratory-reared Anopheles mosquitoes
(Vaughan et al. 1994). This is in contrast to observations on
wild-caught An. gambiae from Burkino-Faso (Lombardi et
al. 1987) and western Kenya (Beier et al. 1990) where
sporozoites failed to enter the salivary glands in 43% and 10%
of infected mosquitoes, respectively. However, when
sporozoites from P. gallinaceum oocysts were injected
into Ae. aegypti female mosquitoes only 6.5 to 10.4% of
inoculated sporozoites invaded the salivary glands.
Interestingly, injected salivary gland sporozoites did not
reinvade the glands (Touray et al. 1992). Recently, a
laboratory study on An. tesselatus mosquitoes infected
with different isolates of P. vivax and P.
falciparum from patients living in Sri Lanka showed that
approximately 15% of mosquito batches in which oocysts
developed failed to produce salivary gland sporozoites
(Gamage-Mendis et al. 1993). This discrepancy between
naturally and laboratory-infected mosquitoes could be
attributed to different environmental conditions or mixed
mosquito populations.
It could be expected sporozoites would elicit a humoral
response in the mosquito by activation of the prophenoloxidase
cascade and as a result be killed by melanization. However,
sporozoites in the hemocoele are rarely seen to be melanized
(Brey 1991). Sporozoites might be protected from mosquito
defense reactions against non-self' by antigen sharing. This
has been demonstrated as a potential mechanism for avoidance
of mosquito defence reactions by microfilariae of Brugia
pahangi (Maier et al. 1987). Immunoreactivity to CSP was
observed on the midgut wall of mosquitoes infected with P.
falciparum (Boulanger et al. 1988) and P. yoelii
(Beaudoin et al. 1990). The latter authors also detected
reactivity on uninfected midguts and suggest the presence of a
common determinant between the parasite and the mosquito.
The migration of sporozoites from oocysts to salivary glands
could be active (in contact with basal lamina), passive (in
suspension) or both. If active, the parasite would use a
gliding motility to move across the basal lamina and reach the
gland. Vanderberg (1974) described circular gliding and
attached waving locomotion and movement in sporozoites of
different species when bovine serum albumin was added to
Medium 199. Sporozoites that move over a substratum in
vitro leave behind trails of CS proteins (Stewart &
Vanderberg 1988, 1991, 1992). Soluble CSP was shown to be
present throughout the hemocoel (Robert et al. 1988). Beier et
al. (1992a) concluded that the release of CSP by sporozoites
is a normal but complex mechanism that they interpreted to be
associated with sporozoite survival in the host, is not site-
specific and it would be regulated in response to background
levels of soluble CSP in the environment (negative feedback
mechanism). This idea is supported by previous results showing
that the parasites cease to synthesize CSP during their
journey through the hemolymph but shedding still occurs
(Posthuma et al. 1988, 1989). There is no evidence yet for
chemotaxis but the congregation of sporozoites in the vicinity
of the glands before invasion may support, according to some
workers, a tactile mechanism (Golenda et al. 1990, Meis et al.
1992b). Several polysacharides have been identified that may
orient protozoa towards or away from a stimulus (Van Houten
1988).
The selective invasion of mosquito salivary glands by malarial
parasites is not well understood. Specific recognition of the
glands was shown in P. knowlesi. Oocysts developed
normally on the gut of An. freeborni but sporozoites
were never found in salivary glands. When glands from the
susceptible vector An. dirus were implanted into the
abdomen, they did become infected (Rosenberg 1985). These
experiments demonstrated that the salivary glands themselves
determined specificity. Although invasion of salivary glands
seems to require specific recognition, the mechanism by which
sporozoites recognize, attach to, and penetrate the glands
remains to be determined.
Sporozoites preferentially invade the medial lobe and the
distal portions of the lateral lobes of the salivary glands
where the salivary duct is not chitinous in Anopheles
species (Sterling et al. 1973, James & Rossignol 1991,
Ponnudurai et al. 1991). Hence, the occurrence of a specific
receptor-mediated invasion by sporozoites is plausible.
Perrone et al. (1986) used lectins to characterize
carbohydrate moieties on the basal lamina of Ae.
aegypti salivary glands. As the median and distal lateral
lobes bound a common lectin, RCA 120, whose substrate is b-D-
galactose, the authors suggested that sugars that bind this
lectin serve as candidate residues to which sporozoites may
attach. In contrast, the sporozoite coat binds some lectins
with very low efficiency (Schulman et al. 1980, Turner &
Gregson 1982). However, in a recent scanning electron
microscopic study Meis et al. (1992b) localized P.
falciparum sporozoites in proximal and distal parts but
were unable to identify any specific regions on the glands
where sporozoites penetrate. These authors also showed sites
where the sporozoites have pierced the basal lamina, which
probably explains the presence of CSP on the basal lamina
induced by shedding during penetration (Posthuma et al. 1989)
and the presence of immunoreactive spots of 1-2 mm on the
surface of infected glands (Golenda et al. 1990). Recently,
Touray et al. (1994) developed an in vivo salivary
gland invasion assay and have found that anti-salivary gland
antibodies, sulfated glycosaminoglycans and some lectins,
particularly Suc-WGA, block invasion of sporozoites. Although
the mechanism of blocking is not yet known, those lectins that
blocked invasion bound to salivary glands but did not bind to
sporozoites. Beier et al. (1992a) and Beier (1993) proposed
that as sporozoites invade the salivary glands, the build-up
of CSP is the signal for sporozoites to halt their active
motility, and thus their release of CSP (down-regulation). The
involvement of CSP and other proteins distinct from CSP such
as PySSP2 (Charoenvit et al. 1987) and PfSSP2 (Robson et al.
1988, Rogers et al. 1992a,b) in binding of sporozoites to the
basal lamina of mosquito salivary glands is speculative. As
they share similarities in their structure they might be
related to this process since it has been suggested region II
of CSP is involved in hepatocyte invasion (Cerami et al.
1992a,b, Pancake et al. 1992).
Rosenberg (1985) and King (1988) have suggested that
sporozoites invade the salivary gland cells using a mechanism
similar to invasion of erythrocytes by merozoites although
there is no evidence of parasitophorous vacuole membrane
formation observed in other stages of the parasite cycle
(Sinden & Strong 1978, Meis & Verhave 1988). Membrane-limited
vacuoles beneath the plasma membrane in An. stephensi
distal salivary gland cells invaded by P. berghei have
been described (Sterling et al. 1973). Penetration could also
involve an unusual intercellular route as suggested by Golenda
et al. (1990) who detected CSP in the region between cells of
the median lobe of the gland. Posthuma et al. (1989) observed
many sporozoites on the basal side of the cells, but also
found trail-like CSP immunoreactivity at the lateral space
between the cells.
After penetration, sporozoites are present in bundles in the
accini of gland cells in both proximal and distal areas. Most
probably, the sporozoites which are present in proximal areas
of the glands are unable to reach the draining duct because of
the chitinized layer in that area whereas distally localized
sporozoites reach the draining duct via the large unchitinized
collecting tube (Meis et al. 1992b). This explanation would
not be valid for Aedes species since the salivary ducts
are lined with chitin and extend the full length of the glands
(James & Rossignol 1991). Penetration by sporozoites could
cause pathological vesiculation and cytoskeletal changes in
salivary gland tissues as the infected cells are often
deformed and swollen (Sterling et al. 1973, Maier et al.
1987). The efficiency of salivary gland invasion is poorly
understood. It has been estimated that the median sporozoite
load in the glands is less than 10,000 in colonized or wild
Anopheles species (Shute 1945, Pringle 1966, Wirtz et
al. 1987, Beier et al. 1991b, Sattabongkot et al. 1991) or
slightly higher (Ponnudurai et al. 1991).
The development of infectivity by the sporozoites appears to
be asynchronous, in some cases taking place in the hemocoele,
while in others not occuring until after they have invaded the
salivary glands. Thus, it seems to be time-dependent rather
than site-dependent (Vanderberg 1975, Daher & Krettli 1980).
It was demonstrated in P. berghei-infected mice that
populations of salivary gland sporozoites were more than
10,000 times as infective to the vertebrate host as
populations of oocyst sporozoites from the same mosquitoes
(Vanderberg 1975). Touray et al. (1992) found that as few as
10-50 salivary gland P. gallinaceum sporozoites were
required to induce infection in chickens compared to 5,000
oocyst sporozoites. Sporozoite infectivity increases with time
during the first week after the invasion of the salivary gland
(Vanderberg 1975). Degeneration of sporozoites is not frequent
in nature as observed by Barber (1936) who studied anophelines
collected in Mediterranean areas. When degeneration occurred
in a salivary gland or a lobe of a gland, in another gland or
lobe the sporozoites were normal or in a different stage of
degeneration.
The number of sporozoites injected into the tissue or
capillary of the vertebrate is very small compared to that
found in the salivary glands. It has been estimated, by
employing different methods, that each bite delivers fewer
than 50 sporozoites with a tendency for most sporozoites to be
ejected in the first droplets of saliva (Vanderberg 1977,
Rosenberg et al. 1990, Ponnudurai et al. 1991, Beier et al.
1991a,b, Beier et al. 1992b, Li et al. 1992). Although a
correlation between salivary gland sporozoite load and
sporozoite inoculum has been reported (Rosenberg et al. 1990)
ejection of sporozoites is probably a random process, more
related to the architecture of the salivary gland duct system
than to the number of sporozoites in this organ (Ponnudurai et
al. 1991).
Some studies reported that P. falciparum-infected
mosquitoes deliver sporozoites in an unpredictable fashion,
sometimes not at all (Ponnudurai et al. 1991), and others
transmit inconsistently (Rickman et al. 1990). Clumping of
sporozoites has been reported in infected salivary glands
(Sterling et al. 1973) and clusters of sporozoites were
detected after delivery when An. stephensi mosquitoes
infected with P. falciparum were allowed to feed
through fresh mouse skin (Ponnudurai et al. 1991). It has been
observed that salivary glands are not depleted of sporozoites
even in vectors that feed up to 15 times (Shute 1945), which
allows infected mosquitoes to remain potentially infectious
for life.
The low sporozoite inoculum and the low entomological
inoculation rates in natural conditions (Mendis et al. 1990a,
Gordon et al. 1991) demonstrates the high efficiency with
which injected sporozoites will develop malaria.
Factors affecting parasite development to the
mosquito
The process of infection of mosquitoes is exceedingly complex
and regulated by a range of factors originating from the
parasite, the vertebrate host and the mosquito vector, and
from the interactions between all three (Sinden 1991). Many of
these and other factors are known to affect fertilization and
subsequent stages of parasite development, thus having great
influence on transmission of the disease. To have an idea,
when sporogonic development of cultured P. falciparum
was evaluated in six species of Anopheles mosquitoes
there was a total loss of approximately 31,600-fold in the
parasite population from macrogametocyte to oocyst stage
(Vaughan et al. 1994).
a) Host location and feeding behaviour -
Of primary importance in malaria transmission is the
proportion of mosquito feeds taken from humans and the
proportion of these feeds taken from infected individuals. In
nature a vector needs to survive longer than the sporogonic
period after taking an infective blood meal; during this
period the mosquito probably takes blood meals every 2-3 days,
depending on its gonotrophic cycle and the availability of
breeding sites (Ponnudurai et al. 1989). Parasitemic hosts
tend to be sick and often less irritated by mosquito feeding.
Additionally, the thrombocytopenia which is commonly
associated with blood-borne parasitic diseases leads to
facilitation of vessel location, resulting in increased
feeding success by mosquitoes on parasitemic hosts (Rossignol
et al. 1985). Salivary glands and saliva contain a whole range
of components with pharmacological activities important for
blood feeding success and subsequent bloodmeal processing,
including anticoagulants, anti-inflammatory, vasodilatory and
immunosuppressive compounds (Ribeiro et al. 1984, 1989,
Ribeiro 1987, Titus & Ribeiro 1990, James & Rossignol 1991).
Rossignol et al. (1984) concluded that the lesions caused by
P. gallinaceum sporozoites in the salivary glands of
Ae. aegypti result in reduced levels of salivary
apyrase. Mosquitoes deficient in salivary apyrase experience
difficulty in locating host blood and engorging; they
therefore probe more often and may attempt to feed on several
hosts in succession (Rossignol et al. 1984, 1986, Ribeiro et
al. 1985). Li et al. (1992) demonstrated that probing time of
P. berghei-infected An. stephensi is not
affected by sporozoite invasion of salivary glands.
Blood meal size may influence infectivity since it determines
the number of gametocytes ingested and therefore subsequent
infection. It has been shown that larger An. dirus
females took larger bloodmeals by artificial feeding with
cultured P. falciparum and developed significantly more
oocysts (Kitthawee et al. 1990). In a field study, Lyimo and
Koella (1992) reported that the proportion of An.
gambiae mosquitoes infected with P. falciparum
during a blood meal was independent of size but the number of
oocysts harboured by infected mosquitoes increased with size
of the mosquito.
b) Gametocyte carriers/transmission blocking immunity -
Malarial infections induce host responses to both asexual and
sexual stage malaria parasites that may modulate gametocyte
infectivity. Great heterogeneity in infectiousness of
different carriers has been noted, with apparently poor
correlation between infectiousness and gametocyte density
(Graves et al.1988). However, it remains unclear whether
symptomatic and asymptomatic asexual infections differ in
their ability to influence gametocyte infectivity. It has been
reported that asymptomatic P. falciparum patients were
more infectious than symptomatic patients (Boyd & Kitchen
1937, Jeffery & Eyles 1955, Muirhead-Thomson 1957, Carter &
Graves 1988) while, in contrast, another report suggested that
asymptomatic and symptomatic P. malariae patients were
equally infective (Young & Burgess 1961).
Specific and non-specific responses are believed to modulate
parasite transmission. The induction of specific immunological
responses to Plasmodium shows marked heterogeneity
within human populations (Mendis et al. 1987, Graves et al.
1988, Targett 1990, Snow et al. 1993). The sexual stages can
induce trasmission blocking immunity (TBI) and effective
targets include antigens identified on the surface of the
macro- and/or microgemetes ( pre-fertilization'antigens) as
well as antigens present on the surface of the gamete, zygote
and ookinete ( post-fertilization' antigens). Antibodies to
pre-fertilization' antigens of P. falciparum such as
Pfs230, Pfs48/50 and Pfs16 have been detected in humans
(Graves et al. 1988, Premawansa et al. 1994, Hogh et al.
1994). A significant association between lacking of
infectivity of P. falciparum gametocyte carriers and
recognition of epitope IIa on Pfs48/50 by antibodies in their
sera has been observed (Graves et al. 1992). Naturally
acquired TBI to P. vivax sexual stage antigens has also
been demonstrated (Mendis et al. 1987, 1990b, Mendis & Carter
1991, Ranawaka et al. 1988, Goonewardene et al. 1990, Gamage-
Mendis et al. 1992) and appears to play a role on transmission
of the disease (de Zoysa et al. 1988). In P. vivax
malaria antibodies, at low concentrations, can also have a
transmission-enhancing effect on infectivity of malarial
parasite to mosquitoes (Mendis et al. 1987, Peiris et al.
1988, Naotunne et al. 1990, Gamage-Mendis et al. 1992). A
recent report described TBI in P. vivax malaria when
antibodies raised against a peptide blocked parasite
development in the mosquito An. tesselatus (Snewin et
al. 1995). As shown in a variety of Plasmodium species,
within the mosquito vector antibody to the pre-fertilization'
antigens may prevent fertilization by any of four mechanisms:
(1) the agglutination of macro/microgametes limiting their
mobility; (2) antibody coating of macro/microgamentes
inhibiting cell-cell recognition; (3) opsonization in the
bloodmeal or (4) complement dependent/independent lysis (Gwadz
1976, Carter & Chen 1976, Carter et al. 1979, 1985, 1990,
Kaushal et al. 1983a,b, Rener et al. 1983, Harte et al. 1985a,
Vermeulen et al. 1985b, Grotendorst et al. 1986, Grotendorst &
Carter 1987, Quakyi et al. 1987, Peiris et al. 1988,
Premawansa et al. 1990). Cellular responses are also involved
in TBI and may have some influence on parasite infectivity
(Harte et al. 1985b, Mendis et al. 1990b, Riley & Greenwood
1990, Goonewardene et al. 1990).
Immunity to `post-fertilization' antigens such as Pfs25,
Pbs21, Pgs25 and Prs25 also plays an important role on
transmission of the disease by suppressing parasite
infectivity at different stages of its development in the
mosquito as demonstrated by many workers in different
Plasmodium species (Grotendorst et al. 1984, Vermeulen
et al. 1985a, Sinden et al. 1987, Winger et al. 1988, Fries et
al. 1989, Carter & Kaushal 1984, Carter et al. 1989a,b, Kaslow
et al. 1991, 1992, 1994b, Foo et al. 1991, Sieber et al. 1991,
Tirawanchai et al. 1991, Duffy et al. 1993, Paton et al. 1993,
Ranawaka et al. 1993, 1994). The mechanisms of blockade could
be the same four described above and/or the antibody may act
by damaging the parasite surface coat (Ponnudurai et al.
1987).
Non-specific responses to the asexual stages are believed to
modulate parasite trasmission (Naotunne et al. 1991,
Kwiatkowski 1992). Numerous non-specific factors may correlate
with changes in gametocyte infectivity. Acute phase reactants
like C-reactive protein (CRP), which are non-specific
indicators of inflammatory activity, are elevated in patients
with P. falciparum malaria (Ree 1971, Naik & Voller
1984, Chagnon et al. 1992). Some cytokines such as interferon
(IFN-g), tumor necrosis factor (TNF-a) and interleukin 6 (IL-
6) are elevated in sera fom patients with P. falciparum
and P. vivax malaria (Grau et al. 1989, Kern et al.
1989, Kwiatkowski et al. 1990, Mendis et al. 1990c,
Karunaweera et al. 1992). Both IFN-g and TNF-a appear to cause
a transient but marked drop in the infectivity of gametocytes
to mosquitoes due to the intraerythrocytic killing of
parasites (Naotunne et al. 1991, Karunaweera et al. 1992).
However, a recent study has shown no elevation in blood levels
of cytokines IL-2, IL-6, TNF-a and IFN-g nor reactive nitrogen
intermediates (Hogh et al. 1994). The authors suggested that
this could be explained by the inability of asymptomatic
gametocyte carriers, unlikely to harbour high asexual
parasitaemias, to promote the responses.
The concept that anti-sexual stage immunity may regulate
infection of the moquito vector by gametocyte-infected
malarial blood has gained considerable support and must be
considered for the development of malaria trasmission blocking
vaccines (Kaslow et al. 1992).
c) Antibodies against sporozoites -
There is evidence that naturally acquired or experimentally
elicited anti-sporozoite antibodies ingested by mosquitoes may
affect the dynamics of the sporogonic development in the
vector. Several studies with P. falciparum-
infected Anopheles species (Vaughan et al. 1988, Beier
et al. 1989, Hollingdale & Rosario 1989) showed that (1)
ingested human CSP antibodies were detected in the blood meal
of field collected mosquitoes up to 36 hr after feeding, (2)
antibodies crossing the midgut into hemocoel persist from 4 to
36 hr post-infection in hemolymph, (3) ingested CSP antibodies
on day 5 after infection bound to developing oocysts, (4)
enhancement of the sporozoite production, (5) ingestion of CSP
antibodies on day 10 after feeding had no effect on oocyst
maturation or sporozoite production, (6) contact between CSP
antibodies and sporozoites in the hemocoel did not block
sporozoite invasion of salivary glands, (7) exposure to CSP
antibody increased sporozoite infectivity and (8) human IgG
antibodies were present on salivary gland sporozoites from
field-collected mosquitoes. It has been recently demonstrated
that antibodies to P. gallinaceum CSP prevent
sporozoites from invading salivary glands of Ae. aegypti
(Warburg et al. 1992). Ponnudurai et al. (1989) did not
find any influence of anti-sporozoite antibodies on the number
of salivary gland sporozoites but concluded that a second
blood meal, with or without antibody, simply functions as a
nutritional stimulus for faster oocyst maturation. However,
when transmission blocking antibodies anti-Pbs21 (a surface
antigen present on the surface of P. berghei
zygote/ookinete) were added to second bloodfeeds at different
stages of parasite development in the mosquito, a significant
reduction in oocyst intensity but no detectable change in
prevalence occurred. Furthermore, at all times anti-Pbs21
reduced sporozoite number in the thorax but highest gland
intensities were obtained when the second bloodfeed was given
on day 4 (Ranawaka et al. 1993). These results were
interpreted as two opposing roles of second bloodfeeds
containing trasmission blocking antibody: (1) inhibition of
parasite development and (2) the supply of nutrients which
permit more sporozoites to be produced by each oocyst. Despite
some controversy these results potentially have significant
implications for natural malaria transmission and for a
possible vaccine development.
d) Anti-mosquito antibodies -
In addition to anti-parasite antibodies, it has been tested
experimentally the effect on malaria trasmission of antibodies
raised against parts of the mosquito which could be included
in a malaria vaccine. Ramasamy and Ramasamy (1990), studying
the P. berghei/An. farauti model, found that
mosquitoes feeding on mice immunised with midgut antigens
exhibited a reduction in mosquito infection rates. Similar
results were reported by Billingsley et al. (1990) using
monoclonal antibodies produced against mosquito midgut tissue
in P. berghei/An. stephensi system.
e) Genetic manipulation of the vector -
Experiments with refractory lines of An. gambiae
(Collins et al. 1986) and studies on the effects of broad
antimicrobial and antiparasitic components, e.g. magainins and
cecropins (Gwadz et al. 1989), showed that it may be feasible
to induce effective disruption in the normal development of
Plasmodium species in the vector by the introduction
and expression of appropriate genes into the mosquito genome.
Two types of useful target genes can be used in transgenic
mosquitoes. First, those that render populations vulnerable to
subsequent control measures, such as insecticide
susceptibility or temperature sensitivity, and second, those
that interrupt disease transmission by replacing vector with
non-vector forms (Crampton et al. 1990, Kidwell & Ribeiro
1992, Crampton 1994). Although many technical, and perhaps
ethical, problems associated with the wild-release of
transgenic insects have yet to be overcome, the potential of
this technology has received greater attention recently (Brey
1991, Coluzzi 1992, Collins 1994, Curtis 1994). Introduction
and expression of genes coding for antibodies against target
antigens present on the ookinete surface into the mosquito
embryos is one of the possibilities to examine the potential
of this technology (Crampton et al. 1993).
f) Anti-malarial drugs -
Sub-therapeutic doses of antimalarial drugs have been reported
to enhance infectivity of Plasmodium species to their
vectors (Shute & Maryon 1954). Additionally, numerous
compounds including chloroquine (Wilkinson et al. 1976),
sulphamethoxazole-trimethroprim (Wilkinson et al. 1973),
pyrimethamine (Shute & Maryon 1951), Fansidar (Carter & Graves
1988) and Berenil (Ono et al. 1993) have been suggested to
induce gametocyte formation but no influence of chloroquine
(Jeffery et al. 1956, Smalley 1977, Chutmongkonkul et al.
1992, Hogh et al. 1994) and Fansidar (Hogh et al. 1994) on
gametocyte infectivity was observed by some investigators. It
was demonstrated that pyrimethamine- and halofantrine-treated
gametocytes of P. falciparum are more infective to
An. stephensi mosquitoes than untreated controls
(Chutmongkonkul et al. 1992). Other studies examined the
effects of some schizontocidal agents on the sporogonic cycle
of P. falciparum and P. berghei in anopheline
mosquitoes (Coleman et al. 1988, do Rosario et al. 1988). It
was found that chloroquine, when fed during late sporogony
(10-12 days post-infection), may increase the vectorial
capacity of some mosquito species. The effects of chloroquine
on the infectivity of chloroquine-sensitive and -resistant
populations of P. yoelii nigeriensis to An.
stephensi mosquitoes were studied by Ichimori et al.
(1990). The results showed an enhancement of infectivity in
sensitive strains but no effect was detected in resistant
clones and sublines. Chloroquine use and the subsequent
development of resistance over the past years is associated
with an increasing human malaria infectiousness (Lines et al.
1991) which may be indirect effects of parasitaemia on the
host. The sporontocidal activity of chloroquine, halofantrine
and pyrimethamine was evaluated by administration to An.
stephensi mosquitoes, either in the first bloodmeal
containing P. falciparum gametocytes from in
vitro cultures, or in the second, parasite-free bloodmeal,
given four days after infection. A sporontocidal effect was
observed only when pyrimethamine was administred with the
infective bloodmeal (Chutmongkonkul et al. 1992). It has been
demonstrated recently an inhibitory action of the anti-
malarial Atovaquone (566C80) against ookinete, oocysts and
sporozoites of Plasmodium berghei in An.
stephensi (Fowler et al.1994, 1995).
Acknowledgments
Amauri Braga Simonetti was supported by a scholarship from
CNPq (Brazil) during his PhD in the Infection and Immunity
Section, Department of Biology at Imperial College of Science,
Technology and Medicine (London, U.K.).
I would like to acknowlegde Professor Robert E Sinden for his
guidance and helpfull advices which made possible this
review.
References
Aikawa M 1971. Plasmodium: The fine structure of
malarial parasites. Exper Parasitol 30: 284-
320.
Aikawa M Atkinson CT, Beaudoin LM, Sedegah M, Charoenvit Y,
Beaudoin R 1990. Localization of CS and Non-CS Antigens in the
Sporogonic Stages of Plasmodium yoelii. Bull
WHO 68: 165-171.
Alano P 1991. Plasmodium sexual stage antigens.
Parasitol Today 7: 199-203.
Alano P, Carter R 1990. Sexual differentiation in malaria
parasites. Ann Rev Microbiol 44: 429-449.
Bafort JM 1971. The biology of rodent malaria with particular
reference to Plasmodium vinkei vinkei Rhodain 1952.
Ann Soc belge de Med Trop 51: 1-204.
Ball G H, Chao J 1957. Development in vitro of isolated
oocysts of Plasmodium relictum. J Parasitol
43: 409-412.
Ball GH, Chao J 1960. In vitro development of the mosquito
phase of Plasmodium relictum. Exp Parasitol
9: 47-55.
Ball GH, Chao J 1961. Infectivity to canaries of sporozoites
of Plasmodium relictum developing in vitro. J
Parasitol 47: 787-790.
Ball GH, Chao J 1976. Use of amino acids by Plasmodium
relictum oocysts in vitro. Exp Parasitol
39: 115-118.
Barber MA 1936. Degeneration of the sporozoites of the
malaria parasite in anopheline mosquitoes in nature and its
relation to the transmission of malaria. Am J Hyg
24: 45-56.
Beaudoin RL, Strome CPA, Tubergen TA 1974. Plasmodium
berghei berghei: ectopic development of the ANKA strain in
Anopheles stephensi. Exp Parasitol 36:
189-201.
Beier JC 1993. Malaria sporozoites: survival, transmission and
disease control. Parasitol Today 9: 210-215.
Beier JC, Davis JR, Vaughan JA, Noden BH, Beier MS 1991a.
Quantitation of Plasmodium falciparum sporozoites
transmitted in vitro by experimentally infected Anopheles
gambiae and Anopheles stephensi. Am J Trop Med
Hyg 44: 564-570.
Beier JC, Onyango FK, Ramadhan M, Koros JK, Asiago C.M, Wirtz
RA, Koech DK, Roberts CR 1991b. Quantitation of malaria
sporozoites in the salivary glands of wild afrotropical
Anopheles. Med Vet Entomol 5: 63-70.
Beier JC, Oster CN, Koros JK, Onyango FK, Githeko AK, Rowton
E, Koech DK, Roberts CR 1989. Effect of human circumsporozoite
antibodies in Plasmodium -infected Anopheles
(Diptera: Culicidae). J Med Entomol 26: 547-
553.
Beier JC, Perkins PV, Koros JK, Onyango FK, Gargan TP, Wirtz
RA, Koech DK, Roberts, CR 1990. Malaria sporozoite detection
by dissection and ELISA to assess infectivity of afrotropical
Anopheles (Diptera: Culicidae). J Med Entomol
27: 377-384.
Beier JC, Vaughan JA, Madani A, Noden, BH 1992a. Plasmodium
falciparum: release of circumsporozoite protein by
sporozoites in the mosquito vector. Exp Parasitol 75:
248-256.
Beier MS, Davis JR, Pumpuni CB, Noden BH, Beier JC 1992b.
Ingestion of Plasmodium falciparum sporozoites during
transmission by anopheline mosquitoes. Am J Trop Med
Hyg 47: 195-200.
Berner R, Rudin W, Hecker H 1983. Peritrophic membranes and
protease activity in the midgut of the malaria mosquito,
Anopheles stephensi (Liston) (Insecta: Diptera) under
normal and experimental conditions. J Ultrastruct Res
83: 195-204.
Billingsley PF 1990. The midgut ultrastructure of
haematophagous insects. Ann Rev Entomol 35:
219-248.
Billingsley PF, Hecker H 1991. Blood digestion in the
mosquito, Anopheles stephensi Liston (Diptera:
Culicidae): activity and distribution of trpsin,
aminopeptidase, and alpha-glucosidase in the midgut. J
Med Entomol 28: 865-871.
Billingsley PF, Rudin W 1992. The role of the mosquito
peritrophic membrane in bloodmeal digestion and infectivity of
Plasmodium species. J Parasitol 78:
430-440.
Billingsley PF, Winger L, Simonetti AB, Sinden REE 1990.
Monoclonal antibdies produced against mosquito midgut tissue.
VII International Congress of Parasitology (COPA), Paris.
Boulanger N, Matile H, Betschart B 1988. Formation of the
circumsporozoite protein of Plasmodium falciparum in
Anopheles stephensi. Acta Trop 45: 55-
65.
Boyd MF, Kitchen SF 1937. On the infectiousness of patients
infected with Plasmodium vivax and Plasmodium
falciparum. Am J Trop Med Hyg 17: 253-
262.
Brey PT 1991. The Taming of the Anopheles - Current
trends in malaria vector research. Res Immunol
142: 712-722.
Briegel H, Lea AO 1975. Relationship between protein and
proteolytic activity in the midgut of mosquitoes. J Ins
Physiol 21: 1597-1604.
Bruce MC, Alano P, Duthie S, Carter R 1990. Commitment of the
malaria parasite Plasmodium falciparum to sexual and
asexual development. Parasitology 100: 191-
200.
Canning EU, Sinden RE 1973. The organisation of the ookinete
and observations on nuclear division in oocysts of
Plasmodium berghei. Parasitology 67: 29-
40.
Carter R, Bushell G, Saul A, Graves PM, Kidson C 1985. Two
apparently nonrepeated epitopes on gametes of Plasmodium
falciparum are targets of transmission-blocking
antibodies. Infect Immun 50: 102-106.
Carter R, Chen DH 1976. Malaria transmission blocked by
immunisation with gamets of the malaria parasite.
Nature 263: 57-60.
Carter R, Graves PM 1988. Gametocytes. In: Malaria.
Principles and practice of malariology, edited by
Wernsdorfer, W.H. and
McGregor, Sir I. Edinburgh: Chuchill Livingstone, p. 253-
306.
Carter R, Graves PM, Creasey A, Byrne K, Read D, Alano P,
Fenton B 1989a. Plasmodium falciparum: an abundant
stage-specific protein expressed during early gametocyte
development. Exp Parasitol 69: 140-149.
Carter R, Graves PM, Keister DB, Quakyi IA 1990. Properties
of epitopes of Pfs 48/45, a target of transmission blocking
monoclonal antibodies, on gametes of different isolates of
Plasmodium falciparum. Paras Immunol 12:
587-603.
Carter R, Graves P M, Quakyi I, Good MF 1989b. Restricted or
absent immune responses in human populations to Plasmodium
falciparum gamete antigens that are targets of malaria
transmission-blocking antibodies. J Exp Med 169:
135-147.
Carter R, Gwadz RW, Mcauliffe FM 1979. Plasmodium
gallinaceum: transmission blocking immunity in chickens.
I. Comparative immunogenicity of gametocyte and gamete-
containing preparations. Exp Parasitol 47:
185-193.
Carter R, Kaushal DC 1984. Characterization of antigens on
mosquito midgut stages of Plasmodium gallinaceum. III.
changes in zygote surface proteins during transformation to
mature ookinete. Mol Biochem Parasitol 13: 235-
241.
Carter R, Kumar N, Quakyi I, Good M, Mendis K, Graves P,
Miller LH 1988. Immunity to sexual stages of malaria
parasites. Progr Allergy 41: 193-214.
Carter R, Nijhout MM 1977. Control of gamete formation
(exflagellation) in malaria parasites. Science
195: 407-409.
Cattani JA 1989. Malaria vaccines: results of human trials and
directions of current research. Exp Med 68: 242-
247.
Cerami C, Frevert V, Sinnis P, Takacs B, Clavijo P, Santos MJ,
Nussenzweig V 1992b. The basolateral domain of the hepatocyte
plasma membrane bears receptors for the CSP of Plasmodium
falciparum sporozoites. Cell 70: 1021-
1033.
Cerami C, Kwakyeberko F, Nusswenzweig V 1992a. Binding of
malarial circumsporozoite protein to sulfatides
Chagnon A, Yao N, Carli P, Paris JF, Marlier S, Pierre C,
Bassiere H 1992. La proteine C-reactive dans l'acces palustre.
Presse Med 21: 217-218.
Charoenvit Y, Leef MF, Yuan LF, Sedegah M, Beaudoin RL 1987.
Characterization of Plasmodium yoelii monoclonal
antibodies directed against stage-specific sporozoite
antigens. Infect Immun 55: 604-608.
Chege GMM, Pumpuni CB, Beier JC 1996. Proteolytic enzyme
activity and Plasmodium falciparum sporogonic
development in three species of Anopheles mosquitoes.
J Parasitol 82: 11-16.
Chutmongkonkul M, Maier WA, Seitz HM 1992. Plasmodium
falciparum: effect of chloroquine, halofantrine and
pyrimethamine on the infectivity of gametocytes for An.
stephensi mosquitoes. Ann Trop Med Parasitol
86: 103-110.
Coleman RE, Vaughan JA, Hayes DO, Hollingdale MR, Do Rosario
VE 1988. Effect of mefloquine and artisinin on the sporogonic
cycle of Plasmodium berghei ANKA in Anopheles
stephensi mosquitoes. Acta Leid 57: 61-
74.
Collins FH 1994. Prospects for malaria control through genetic
manipulation of its vectors. Parasitol Today 10:
370-371.
Collins FH, Sakai RK, Vernick KD, Paskewitz S, Seeley DC,
Miller LH, Collins WE, Campbell CC, Gwadz RW 1986. Genetic
selection of a Plasmodium refractory strain of the
malaria vector Anopheles gambiae. Science
234: 607-610.
Coluzzi M 1992. Malaria vector analysis and control.
Parasitol Today 8: 113-118.
Cornelissen AWCA 1988. Sex determination and sex
differentiation in malaria parasites. Biol Rev
63: 379-394.
Crampton JM 1994. Molecular studies of insect vectors of
malaria. Adv Parasitol 34: 1-31.
Crampton JM, Galler R, Sinden RE, Crisanti A 1993. La lutte
genetique contre les moustiques. La Recherche
259: 1218-1219.
Crampton JM, Morris A, Lycett G, Warren A, Eggleston P 1990.
Transgenic mosquitoes: a future vector control strategy?
Parasitol Today 6: 31-36.
Curtis CF 1994. The case for malaria cntrol by genetic
manipulation of its vectors. Parasitol Today 10:
371-374.
Daher VR, Krettli AU 1980. Infectivity of Plasmodium
gallinaceum sporozoites from oocysts. J Protozool
27: 440-442.
Davies EE 1974. Ultrastructural studies on the early ookinete
stage of Plasmodium berghei nigeriensis and its
transformation into an oocyst. Ann Trop Med Parasitol
68: 283-290.
Dearsly AL, Sinden RE, Self IA 1990. Sexual development in
malarial parasites - Gametocyte production, fertility and
infectivity to the mosquito vector. Parasitology
100: 359-368.
De Zoysa APK, Herath PRJ, Abhywrana A, Padmalal UKGK, Mendis
KN 1988. Modulation of human malaria transmission by anti-
gamete transmission blocking immmunity. Trans R Soc Trop
Med Hyg 82: 548-553.
Do Rosario .E, Vaughan JA, Murphy M, Harrod V, Coleman R 1988.
Effect of chloroquine on the sporogonic cycle of Plasmodium
falciparum and Plasmodium berghei in anopheline
mosquitoes. Acta Leid 57: 53-60.
Duffy P, Pimenta P, Kaslow DC 1993. Pgs28 belongs to a family
of epidermal growth factor-like antigens that are targets of
malaria transmission-blocking antibodies. J Exp Med
177: 505-510.
Duncan D, Eades J, Julian SR, Micks D 1960. Electron
microscopic observations on malarial oocysts (Plasmodium
relictum). J Protozool 7: 18-26.
Eyles DE 1952. Studies on Plasmodium gallinaceum II.
Factors in the blood of the vertebrate host influencing
mosquito infection. Am J Trop Med Hyg 5: 276-
290.
Feldmann AM, Billingsley PF, Savelkoul A 1990. Blood meal
digestion by strains of Anopheles stephensi Liston
(Diptera: Culicidae) of differing susceptibility to
Plasmodium falciparum. Parasitology 101:
193-200.
Feldmann AM, Ponnudurai T 1989. Selection of Anopheles
stephensi for refractoriness and susceptibility to
Plasmodium falciparum. Med Vet Entomol
3: 41-52.
Foo A, Carter R, Lambros C, Graves P, Quakyi I, Targett GAT,
Ponnudurai T, Lewis GE 1991. Conserved and variant epitopes of
target antigens of transmission-blocking antibodies among
isolates of Plasmodium falciparum from Malaysia. Am
J Trop Med Hyg 44: 623-631.
Fowler RE, Billingsley PF, Pudney M, Sinden RE 1994.
Inhibitoy action of the anti-malarial Atovaquone (566C80)
against Plasmodium berghei ANKA in the mosquito Anopheles
stephensi. Parasitology 108: 383-388.
Fowler RE, Sinden RE, Pudney M 1995. Inhibitory activity of
the anti-malarial Atovaquone (566C80) against ookinete,
oocysts and sporozoites of Plasmodium berghei. J
Parasitology 8: 452-458.
Freier J E, Friedman S 1987. Effect of Plasmodium
gallinaceum infection on the mortality and body weight of
Aedes aegypti (Diptera : Culicidae). J Med
Entomol 24: 6-10.
Freyvogel TA 1966. Shape, movement in situ and locomotion of
plasmodial ookinetes. Acta Trop 23: 201-221.
Fries HCW, Lamers MBAC, Smits MA, Ponnudurai T, Meuwissen JHET
1989. Characterization of epitopes on the 25 kiloDalton
protein of the macrogametes/zygotes of Plasmodium
falciparum. Parasite Immunol 11: 31-45.
Gamage-Mendis AC, Rajakaruna J, Carter R, Mendis KN 1992.
Transmission blocking immunity to human Plasmodium
vivax malaria in an endemic population in Kataragama, Sri-
Lanka. Parasite Immunol 14: 385-396.
Gamage-Mendis AC, Rajakaruna J, Weerasinghe S, Mendis C,
Carter R, Mendis KN 1993. Infectivity of Plasmodium
vivax and Plasmodium falciparum to Anopheles
tessellatus; relationship between oocyst and sporozoite
development. Trans R Soc Trop Med Hyg 87: 3-
6.
Garnham PCC 1966. Malaria parasites and other
haemosporidia. Blackwell Scientific Publications, Oxford,
1114 pp.
Garnham PCC, Bird RG, Baker JR 1962. Electron microscopic
studies of motile stages of malarial parasites III. The
ookinetes of Haemamoeba and Plasmodium. Trans
R Soc Trop Med Hyg 56: 116-120.
Garnhm PCC, Bird RG, Baker JR, Desser SS, El-Nahal HMS 1969.
Electron microscopic studies on the motile stages of malaria
parasites VI. The ookinete of Plasmodium berghei yoelii
and its transformation into an early oocyst. Trans R Soc
Trop Med Hyg 63: 187-194.
Gass RF 1977. Influences of blood digestion on the development
of Plasmodium gallinaceum (Brumpt) in the midgut of
Aedes aegypti (L.). Acta Trop 34: 127-
140.
Gass RF, Yeates RA 1979. In vitro damage to cultured
ookinetes of Plasmodium gallinaceum by digestive
proteinases from susceptible Aedes aegypti. Acta
Trop 36: 243-252.
Golenda CF, Starkweather WH, Wirtz RA 1990. The distribution
of circumsporozoite protein (Cs) in Anopheles stephensi
mosquitoes infected with Plasmodium falciparum malaria.
J Histochem Cytochem 38: 475-481.
Good MF 1991a. Towards the development of the ideal malaria
vaccine - A decade of progress in a difficult field. Med
J Austr 154: 284-289.
Good MF 1991b. The implications for malaria vaccine programs
if memory T-cells from non-exposed humans can respond to
malaria antigens. Curr Op Immunol 3: 496-
502.
Good MF 1992. A malaria vaccine strategy based on the
induction of cellular immunity. Immunol Today 13: 126-
129.
Goonewardene R, Carter R, Gamage CP, Delgiudice G, David PH,
Howie S, Mendis KN 1990. Human T cell proliferative responses
to Plasmodium vivax antigens: evidence of
immunosuppression following prolonged exposure to endemic
malaria. Europ J Immunol 20: 1387-1391.
Gordon DM, Davis DR, Lee M, Lambros C, Harrison BA, Samuel R,
Campbell GH, Jeghathesan M, Selvarajan K, Lewis GE 1991.
Significance of circumsporozoite-specific antibody in the
natural transmission of Plasmodium falciparum, Plasmodium
vivax, and Plasmodium malariae in an aboriginal
(Orang-Asli) population of central peninsular Malaysia. Am
J Trop Med Hyg 45: 49-56.
Graf R, Briegel H 1982. Comparison between aminopeptidase and
trypsin activity in blood-fed females of Aedes
aegypti. Rev Suis Zool 89: 845-850.
Graf R, Raikhel AS, Brown MR, Lea AO, Briegel H 1986. Mosquito
trypsin: immunocytochemical localization in the midgut of
blood-fed Aedes aegypti (L.). Cell Tis Res
245: 19-27.
Grau GE, Taylor TE, Molyneux ME, Wirima JJ, Vassali P, Hommel
M, Lambert PH 1989. Tumor necrosis factor and disease severity
in children with falciparum malaria. N Engl J Med
320: 1586-1591.
Graves PM, Carter R, Burkot TR, Quakyi IA, Kumar N 1988.
Antibodies to Plasmodium falciparum gamete surface
antigens in Papua New Guinea sera. Paras Immunol 10:
209-218.
Graves PM, Doubrovsky A, Satabongkot J, Battistutta D 1992.
Human antibody responses to epitopes on the Plasmodium
falciparum gametocyte antigen PFS48/45 and their
relationship to infectivity of gametocyte carriers. Am J
Trop Med Hyg 46: 711-719.
Grotendorst CA, Carter, R 1987. Complement effects on the
infectivity of Plasmodium gallinaceum to Aedes
aegypti mosquitoes II. changes in sensitivity to
complement-like factors during zygote development. J
Parasitol 73: 980-984.
Grotendorst CA, Carter R, Rosenberg R, Koontz L 1986.
Complement effects on the infectivity of Plasmodium
gallinaceum to Aedes aegypti mosquitoes I.
Resistance of zygotes to the alternative pathway of
complement. J Immunol 136: 4270-4274.
Grotendorst CA, Kumar N, Carter R, Kaushal DC 1984. A surface
protein expressed during transformation of zygotes of
Plasmodium gallinaceum is a target of transmission-
blocking antibodies. Infec Immun 45: 775-777.
Gwadz W 1976. Malaria: successful immunization against the
sexual stages of Plasmodium gallinaceum. Science
193: 1150-1151.
Gwadz RW, Kaslow D, Lee J-Y, Maloy WL, Zasloff M, Miller LH
1989. Effects of magainins and cercropins on the sporogonic
development of malaria parasites in mosquitoes. Infec
Immun 57: 2628-2633.
Hamilton AJ, Davies CS, Sinden RE 1988. Expression of the
circumsporozoite proteins revealed in situ in the mosquito
stages of Plasmodium berghei by the Lowicryl-immunogold
technique. Parasitology 96: 273-280.
Harte PG, Rogers N, Targett GAT 1985a. Vaccination with
purified microgamete antigens prevents transmission of rodent
malaria. Nature 316: 258-259.
Harte PG, Rogers NC, Targett GAT 1985b. Monoclonal anti-
gamete antibodies prevent transmission of murine malaria.
Paras Immunol 7: 607-615.
Hawking F, Wilson ME, Gammage K 1971. Evidence for the cyclic
development and short-lived maturity in the gametocytes of
Plasmodium falciparum. Trans R Soc Trop Med
Hyg 65: 549-559.
Hecker H 1977. Structure and function of midgut epithelial
cells in Culicidae mosquitoes (Insecta, Diptera).
Cell Tis Res 184: 321-341.
Hollingdale MR, do Rosario V 1989. Malaria transmission-
enhancing activity in mosquitoes by mammalaian host anti-
sporozoite antibodies. Exp Parasitol 68: 365-
368.
Hommel M 1991. Steps towards a malaria vaccine. Res
Immunol 142: 618-631.
Houk EJ, Hardy JL, Chiles RE 1986. Mesenteronal epithelial
cellsurface charge of the mosquito, Culex
tarsalis Coquillett. Binding of colloidal iron
hydroxide, native ferritin and cationised ferritin. J
Submicroscop Cytol 18: 385-396.
Howells RE, Davies EE 1971. Nuclear division in the oocyst of
Plasmodium berghei. Ann Trop Med Hyg 65:
451-459.
Huber M, Cabib E, Miller LH 1991. Malaria parasite chitinase
and penetration of the mosquito peritrophic membrane. Proc
Natl Acad Sci USA 88: 2807-2810.
Ichimori K, Curtis CF, Targett GAT 1990. The effects of
chloroquine on the infectivity of chloroquine-sensitive and -
resistant populations of Plasmodium yoelii nigeriensis
to mosquitoes. Parasitology 100: 377-381.
Iinselburg J 1983. Gametocyte formation by the progeny of
single Plasmodium falciparum schizonts. J
Parasitol 69: 584-591.
James AA, Rossignol PA 1991. Mosquito salivary glands -
Parasitological and molecular aspects. Parasitol Today
7: 267-271.
Jeffery GM, Eyles DE 1955. Infectivity to mosquitoes of
Plasmodium falciparum as related to gametocyte density
and duration of infection. Am J Trop Med Hyg 4:
781-789.
Jeffery GM, Young MD, Eyles DE 1956. The treatment of
Plasmodium falciparum infection with chloroquine, with
a note on infectivity to mosquitoes of primaquine-and
pyrimethamine-treated cases. Am J Hyg 64: 1-
11.
Karunaweera ND, Grau GE, Gamage P, Carter R, Mendis KN 1992.
Dynamics of fever and serum levels of tumor necrosis factor
are closely associated during clinical paroxysms in
Plasmodium vivax malaria. Proc Natl Acad Sci USA
89: 3200-3203.
Kaslow DC, Bathurst IC, Barr PJ 1992. Malaria trasmission-
blocking vaccines. Trends Biotechnol 10: 388-
391.
Kaslow DC, Bathurst IC, Lensen T, Ponnudurai T, Barr PJ,
Keister DB 1994. Saccharomyces cerevisiae recombinant Pfs25
adsorbed to alum elecits antibodies that block transmission of
Plasmodium falciparum. Infec Immun 62: 5576-5580.
Kaslow DC, Isaacs SN, Quakyi IA, Gwadz RW, Moss B, Keister DB
1991. Induction of Plasmodium falciparum transmission
blocking antibodies by recombinant vaccinia virus.
Science 252: 1310-1313.
Kaslow DC, Nussenzweig V, Miller L 1994a. Meeting on parasites
and the invertebrate vector. Mem Inst Oswaldo
Cruz 89: 279-295.
Kaslow DC, Quakyi IA, Syin C, Raum MG, Keister DB, Coligan
JE, McCutchan TF, Miller LH 1988. A vaccine candidate from
the sexual stage of human malaria that contains EGF-like
domains. Nature 333: 74-76.
Kaushal DC, Carter R 1984. Caracterization of antigens on the
mosquito midgut stages of Plasmodium gallinaceum . II.
Comparison of surface antigens of male and female gametes and
zygotes. Mol Biochem Parasitol 11: 145-156.
Kaushal DC, Carter R, Howard RJ, Macauliffe FM 1983a.
Characterization of antigens on mosquito midgut stages of
Plasmodium gallinaceum I. Zygote surface antigens.
Mol Biochem Parasitol 8: 53-69.
Kaushal DC, Carter R, Rener J, Grotendorst CA, Miller LH,
Howard RJ 1983b. Monoclonal antibodies against surface
determinants on gametes of Plasmodium gallinaceum block
transmission of malaria prasites to mosquitoes. J
Immunol 131: 2557-2562.
Kawamoto F, Alejo Blanco R, Fleck SL, Sinden RE 1991.
Plasmodium berghei - Ionic regulation and the induction
of gametogenesis. Exp Parasitol 72: 33-42.
Kern P, Hemmer CJ, Van Damme J, Gruss HJ, Dietrich M 1989.
Elevated tumor necrosis factor-alfa and interleukin-6 serum
levels as markers for complicated Plasmodium falciparum
malaria. Am J Med 87: 139-143.
Kidwell MG, Ribeiro JMC 1992. Can transposable elements be
used to drive disease refractoriness genes into vector
populations? Parasitol Today 8: 325-329.
King C 1988. Cell motility of sporozoan protozoa.
Parasitol Today 4: 315-319.
Kitthawee S, Edman JD, Sattabongkot J 1990. Evaluation of
survival potential and malaria susceptibility among different
size classes of laboratory-reared Anopheles dirus.
Am J Trop Med Hyg 43: 328-332.
Kumar N, Carter R 1984. Biosynthesis of the target antigens of
antibodies blocking transmission of Plasmodium
falciparum. Mol Biochem Parasitol 13: 333-
342.
Kumar N, Carter R 1985. Biosynthesis of two stage-specific
membrane proteins during transformation of Plasmodium
gallinaceum zygotes into ookinetes. Mol Biochem
Parasitol 14: 127-139.
Kwiatowski D 1992. Malaria: becoming more specific about non-
specific immunity. Curr Op Immunol 4: 425-
431.
Kwiatowski D, Hill AVS, Sambou I, Twumasi P, Castracane J,
Manogue KR, Cerami A, Brewster DR, Greenwood BM 1990. TNF
concentrations in fatal cerebral, non-fatal cerebral, and
uncomplicated Plasmodium falciparum malaria.
Lancet ii: 1201-1204.
Lensen AHW, van Gemert GJA, Bolmer MG, Meis JFGM, Kaslow D,
Meuwissen JHE Th, Ponnudurai T 1993. Transmission blocking
antibody of the Plasmodium falciparum zygote/ookinete
surface protein Pfs25 also influences sporozoite development.
Parasite Immunol 14: 471-479.
Li X, Sina B, Rossignol PA 1992. Probing behaviour and
sporozoite delivery by Anopheles stephensi infected
with Plasmodium berghei. Med Vet Entomol 6:
57-61.
Lines J.D, Wilkes TJ, Lyimo EO 1991. Human malaria
infectiousness measured by age-specific sporozoite rates in
An gambiae in Tanzania. Parasitology 102:
167-177.
Lombardi S, Esposito F, Zavala F, Lamizana L, Rossi P,
Sabatinelli G, Nussenzweig R, Coluzzi M 1987. Detection and
anatomical localization of Plasmodium falciparum
circumsporozoite protein and sporozoites in the afrotropical
malaria vector Anopheles gambiae S.L. Am J Trop Med
Hyg 37: 491-494.
Lyimo EO, Koella JC 1992. Relatiosnship between body size of
adult Anopheles gambiae s.l. and infection with the
malaria parasite Plasmodium falciparum.
Parasitology 104: 233-237.
Mack SR, Samuels S, Vanderberg JP 1979. Hemolymph of
Anopheles stephensi from uninfected and Plasmodium
berghei-infected mosquitoes. 2. Free amino acids. J
Parasitol 65: 130-136.
Mack SR, Vanderberg JP 1978. Hemolymph of Anopheles
stephensi from noninfected and Plasmodium berghei-
infected mosquitoes. 1. Collection procedure and physical
characteristics. J Parasitol 64: 918-923.
Maier WA, Becker-Feldman H, Seitz HM 1987. Pathology of
malaria infected mosquitoes. Parasitol Today 3:
216-218.
Martin SK, Miller LH, Nijhout MM, Carter R 1978. Plasmodium
gallinaceum: induction of male gametocyte exflagellation
by
phosphdiesterase inhibitors. Exp Parasitol 44:
239-242.
Mehlhorn H, Peters W, Haberkorn A 1980. The formation of
ookinetes and oocysts in Plasmodium gallinaceum
(Haemosporidia) and considerations on phylogenetic
relationships between Haemosporidia, Piroplasmida and other
Coccidia. Protistologica 16: 135-154.
Meis JFGM, Croes H, Mons B, Vanbelkum A, Ponnudurai T 1992a.
Localization of circumsporo-zoite protein in the sporogonic
stages of Plasmodium vivax. Parasitol Res
78: 165-167.
Meis JFGM, Ponnudurai T 1987. Ultrastructural studies on the
interaction of Plasmodium falciparum ookinetes with the
midgut epithelium of Anopheles stephensi mosquitoes.
Parasitol Res
73: 500-506.
Meis JFGM, Pool G, Vangemert GJ, Lensen AHW, Ponnudurai T,
Meuwissen JHET 1989. Plasmodium falciparum ookinetes
migrate intercellularly through Anopheles stephensi
midgut epithelium. Parasitol Res 76: 13-19.
Meis JFGM, Verhave JP 1988. Exoerythrocytic development of
malarial parasites. Adv Parasitol 27: 1-66.
Meis JFGM, Wismans PGP, Jap PHK, Lensen AHW, Ponnudurai T
1992b. A scanning electron microscopic study of the sporogonic
development of Plasmodium falciparum in Anopheles
stephensi. Acta Trop 50: 227-236.
Mendis KN, Carter R 1991. Transmission blocking immunity may
provide clues that antimalarial immunity is largely T-
independent. Res Immunol 142: 687-690.
Mendis KN, David PH, Carter R 1990b. Human immune responses
against sexual stages of malaria parasites: considerations for
malaria vaccines. Intern J Parasitol 20: 497-
502.
Mendis, KN, Ezoysa APK, Abhayawardena TA, Carter R, Herath
PRJ, Mendis G 1990a. Characteristics of malaria transmission
in Kataragama, Sri-Lanka - A focus for immuno-epidemiological
studies. Am J Trop Med Hyg 42: 298-308.
Mendis KN, Munesinghe YD, DeSilva YNY, Keragalla I, Carter R
1987. Malaria transmission-blocking immunity induced by
natural infections of Plasmodium vivax in humans.
Infec Immun 55: 369-372.
Mendis KN, Naotunne TD, Karunaweera ND, Delgiudice G, Grau GE,
Carter R 1990c. Anti-parasite effects of cytokines in malaria.
Immunol Lett 25: 217-220.
Mitchell GH 1989. An update on candidate malaria vaccines.
Parasitology 98: S29-S47.
Mons B 1985. Induction of sexual differentiation in malaria.
Parasitol Today 1: 87-89.
Mons B 1986. Intraerythrocytic differentiation of
Plasmodium berghei. Acta Leid 54: 1-
83.
Mons B, Janse CJ, Boorsma EG 1985. Synchronized erythrocytic
schizogony and gametocytogenesis of Plasmodium berghei
in vivo and in vitro. Parasitology 91: 423-430.
Muirhead-Thomson RC 1957. The malarial infectivity of an
African village population to mosquitoes (Anopheles
gambiae). A random xenodiagnostic survey. Am J Trop
Med Hyg 6: 971-979.
Nagasawa H, Aikawa M, Procell PM, Campbell GH, Collins WE,
Campbell CC 1988. Plasmodium malariae: distribution of
circumsporozoite protein in the oocyst and salivary gland
sporozoite. Exp Parasitol 66: 27-34.
Nagasawa H, Procell PM, Atkinson CT, Campbell GH, Collins WE,
Aikawa M 1987. Localization of circumsporozoite protein of
Plasmodium ovale in midgut oocysts. Infec Immun
55: 2928-2932.
Naik P, Voller A 1984. Serum C-reactive protein levels and
falciparum malaria. Trans R Soc Trop Med Hyg 78: 812-
813.
Naotunne TD, Karunaweera ND, Delgiudice G, Kularatne MU, Grau
GE, Carter R, Mendis KN 1991. Cytokines kill malaria
parasites during infection crisis - Extracellular
complementary factors are essential. J Exp Med
173: 523-529.
Naotunne TD, Rathnayake KDL, Jayasinghe A, Carter R, Mendis
KN 1990. Plasmodium cynomolgi - Serum-mediated blocking
and enhancement of infectivity to mosquitoes during infections
in the natural host, Macaca sinica. Exp
Parasitol 71: 305-313.
Nijhout MM 1979. Plasmodium gallinaceum: exflagellation
stimulated by a mosquito factor. Exp Parasitol
48: 75-80.
Nijhout MM, Carter R 1978. Gamete development in malarial
parasites: bicarbonate-dependent stimulation by pH in vitro.
Parasitology 76: 39-53.
Nussenzweig RS 1990. Recombinant proteins and synthetic
peptides as malaria vaccine candidates. Can J Microbiol
36: 817-820.
Nussenzweig V, Nussenzweig RS 1989. Rationale for the
development of an engineered sporozoite malaria vaccine.
Adv Immunol 45: 283-334.
Ono T, Onhnishe Y, Nagamune K, Kano M 1993. Gametocytogenesis
induction by Berenil in cultured Plsmodium falciparum.
Exp Parasitol 77: 74-78.
Pancake SJ, Holt GD, Mellouk S, Hoffman SL 1992. Malaria
sporozoites and circumsporozoite proteins bind specifically to
sulfated glycoconjugates. J Cell Biol 117:
1351-1357.
Paskewitz SM, Brown MR, Collins FH, Lea AO 1989.
Ultrastructural localization of phenoloxidase in the midgut of
refractory Anopheles gambiae and association of the
enzyme with encapsulated Plasmodium cynomolgi. J
Parasitol 75: 594-600.
Paton MG, Barker GC, Matsuoka H, Ramesar J, Janse CJ, Waters
AP, Sinden RE 1993. Structure and expression of a post-
transcriptionally regulated malaria gene enconding a surface
protein from the sexual stages of Plasmodium berghei.
Mol Biochem Parasitol 59: 263-276.
Peiris JSM, Premawansa S, Ranawaka MBR, Udagama PV, Munasonghe
YD, Nanayakkara MV, Gamage CP, Carter R, David PH, Mendis KN
1988. Monoclonal and polyclonal antibodies both block and
enhance transmission of human Plasmodium vivax malaria.
Am J Trop Med Hyg 39: 26-32.
Perrone JB, DeMaio J, Spielman A 1986. Regions of mosquito
salivary glands distinguished by surface lectin-binding
characteristics. Insect Biochem 16: 313-318.
Perrone JB, Spielman A 1988. Time and site of assembly of the
peritrophic membrane of the mosquito Aedes aegypti.
Cell Tiss Res 252: 473-478.
Phillips RS 1992. Vaccination against malaria.
Immunobiology 184: 240-262.
Ponnudurai T, Billingsley PF, Rudin W 1988. Differential
infectivity of Plasmodium for mosquitoes. Parasitol
Today 4: 319-321.
Ponnudurai T, Lensen AHW, Van Gemert, GJA, Bensink MPE, Bolmer
M, Meuwissen JHET 1989. Infectivity of cultured Plasmodium
falciparum gametocytes to mosquitoes. Parasitology
98: 165-173.
Ponnudurai T, Lensen AHW, Van Gemert GJA, Bolmer MG, Meuwissen
JHET 1991. Feeding behaviour and sporozoite ejection by
infected Anopheles stephensi. Trans R Soc Trop Med
Hyg 85: 175-180.
Posthuma G, Meis FGM, Verhave JP, Hollingdale MR, Ponnudurai
T, Meuwissen JHET 1988. Localization of circumsporozoite
protein of the malaria parasite Plasmodium falciparum
during sporogony in Anopheles stephensi midguts using
immunoelectron-microscopy of ultrathin cryosections. Eur J
Cell Biol 46: 18-24.
Posthuma G, Meis JFGM, Verhave JP, Gigengak S, Hollingdale MR,
Ponnudurai T, Genze HJ 1989. Immunogold determination of
Plasmodium falciparum circumsporozoite protein in
Anopheles stephensi salivary gland cells. Eur J
Cell Biol 49: 66-72.
Premawansa S, Gamage-Mendis A, Perera L, Begarnie S, Mendis K,
Carter R 1994. Plasmodium falciparum malaria
trasmission-blocking immunity under conditions of low
endemicity as in Sri Lanka. Parasite Immunol 16:
35-42.
Premawansa S, Peiris JSM, Perera KLRL, Ariyaratne G, Carter R,
Mendis KN 1990. Target antigens of transmission blocking
immunity of Plasmodium vivax malaria - Characterization
and polymorphism in natural parasite isolates. J
Immunol 144: 4376-4383.
Pringle G 1965. A count of the sporozoites in an oocyst of
Plasmodium falciparum. Trans R Soc Trop Med Hyg
59: 289-290.
Pringle G 1966. A quantitative study of naturally acquired
malaria infections in Anopheles gambiae and
Anopheles funestus in a highly malarious area of east
Africa. Trans R Soc Trop Med Hyg 60: 626-
632.
Quakyi IA, Carter R, Rener J, Kumar N, Good MF, Miller LH
1987. The 230-kDa gamete surface protein of Plasmodium
falciparum is also a target for transmission-blocking
antibodies. J Immunol 139: 4213-4217
Ramasamy M S, Ramasamy R 1990. Effect of anti-mosquito
antibodies on the infectivity of the rodent malaria parasite
Plasmodium berghei to Anopheles farauti. Med
Vet Entomol 4: 161-166.
Ranawaka G, Alejo Blanco R, Sinden RE 1993. The effect of
transmission-blocking antibody ingested in primary and
secondary bloodfeeds, upon the development of Plasmodium
berghei in the mosquito vector. Parasitology
107: 225-231.
Ranawaka GRR, Fleck SL, Alejo Blanco AR, Sinden RE 1994.
Characterization of the modes of action of anti-Pbs21 malaria
transmission-blocking immunity: ookinete to oocyst
differentiation in vivo. Parasitology
109: 403-411.
Ranawaka MB, Munesinghe YD, De Silva DMR, Carter R, Mendis KN
1988. Boosting of transmission-blocking immunity during
natural Plasmodium vivax infections in humans depends
upon frequent reinfection. Infec Immun 56:
1820-1824.
Ree GH 1971. C-reactive protein in Gambian africans with
special reference to P. falciparum malaria. Trans R.
Soc Trop Med Hyg 65: 574-580.
Rener J, Graves PM, Carter R, Williams JL, Burkot T 1983.
Target antigens of transmission-blocking immunity on gametes
of Plasmodium falciparum. J Exp Med 158:
971-976.
Ribeiro JMC 1987. Role of saliva in blood-feeding by
arthropods. Ann Rev Entomol 32: 463-478.
Ribeiro JMC, Modi GB, Tesh RB 1989. Salivary apirase activity
of some old world phlebotomine sand flies. Insect
Biochem 19: 409-412.
Ribeiro MC, Rossignol PA, Spielman A 1984. Role of mosquito
saliva in blood vessel location. J Exp Biol
108: 1-9.
Ribeiro JMC, Rossignol PA, Spielman A 1985. Salivary gland
apyrase activity determines probing time in Anopheline
mosquitoes. J Insect Physiol 31: 689-692.
Rickman LS, Jones TR, Long GW, Paparello S, Schneider I, Paul
CF, Beaudoin RL, Hoffman SL 1990. Plasmodium
falciparum-infected Anopheles stephensi
inconsistently transmit malaria to humans. Am J Trop Med
Hyg 43: 441-445.
Riley E, Greenwood B 1990. Measuring cellular immune responses
to malaria antigens in endemic populations - Epidemiological,
parasitological and physiological factors which influence in
vitro assays. Immunol Lett 25: 221-229.
Robert V, Verhave JP, Ponnudurai T, Louwe L, Scholtens P,
Carnevale P 1988. Study of the distribution of
circumsporozoite antigen in Anopheles gambiae infected
with Plasmodium falciparum, using the enzyme-linked
immunosorbent assay. Trans R Soc Trop Med Hyg
82: 389-391.
Robson KJH, Hall JRS, Jennings MW, Harris TJR, Marsh K,
Newbold CI, Tate VE, Weatherhall DJ 1988. A highly conserved
amino-acid sequence in thrombospondin, properdin and in
proteins from sporozoites and blood stages of a human malaria
parasite. Nature 335: 79-82.
Rogers WO, Malik A, Mellouk S, Nakamura K, Rogers MD, Szarfman
A, Gordon DM, Nussler AK, Aikawa M, Hoffman SL 1992b.
Characterization of Plasmodium falciparum sporozoite
surface protein 2. Proc Natl Acad Sci USA 89: 9176-
9180.
Rogers WO, Rogers MD, Hedstrom RC, Hoffman SL 1992a.
Characterization of the gene encoding sporozoite surface
protein-2, a protective Plasmodium yoelii sporozoite
antigen. Mol Biochem Parasitol 53: 45-51.
Rosenberg R 1985. Inability of Plasmodium knowlesi
sporozoites to invade Anopheles freeborni salivary
glands. Am J Trop Med Hyg 34: 687-691.
Rosenberg R, Koontz LC 1984. Plasmodium gallinaceum:
density dependent limits on infectivity to Aedes
aegypti. Exp Parasitol 57: 234-238.
Rosenberg R, Koontz LC, Alston K, Friedman FK 1984.
Plasmodium gallinaceum: erythrocyte factor essential
for zygote infection of Aedes aegypti. Exp
Parasitol 57: 158-164.
Rosenberg R, Rungsiwongse J 1991. The number of sporozoites
produced by individual malaria oocysts. Am J Trop Med
Hyg 45: 574-577.
Rosenberg R, Wirtz RA, Scneider I, Burge R 1990. An
estimation of the number of malaria sporozoites ejected by a
feeding mosquito. Trans R Soc Trop Med Hyg 84:
209-212.
Rossignol PA, Ribeiro JMC, Jungery M, Turell MJ, Spielman A,
Bailey CL 1985. Enhanced mosquito blood-finding success on
parasitaemic hosts: evidence for vector-parasite mutualism.
Proc Natl Acad Sci USA 82: 7725-7727.
Rossignol PA, Ribeiro JMC, Spielman A 1984. Increased
intradermal probing time in sporozoite-infected mosquitoes.
Am J Trop Med Hyg 33: 17-20.
Rossignol PA, Ribeiro JMC, Spielman A 1986. Increased biting
rate and reduced fertility in sporozoite-infected mosquitoes.
Am J Trop Med Hyg 35: 277-279.
Roush RT 1993. Occurrence, genetics and management of
insecticide resistance. Parasitol Today 9: 174-
179.
Rowland M, Boersma E 1988. Changes in the spontaneous flight
activity of the mosquito Anopheles stephensi by
parasitization with the rodent malaria Plasmodium
yoelii. Parasitology 97: 221-227.
Rudin W, Billingsley PF, Saladin S 1991. The fate of
Plasmodium gallinaceum in Anopheles stephensi
Liston and possible barriers to transmission. Ann Soc Belg
Med Trop 71: 167-177.
Rudin W, Hecker H 1989. Lectin-binding sites in the midgut of
the mosquitoes Anopheles stephensi Liston and Aedes
aegypti L. (Diptera: Culicidae0. Parasitol Res
75: 268-279.
Rutledge LC, Gould DJ, Tantichareon B 1969. Factors affecting
the infection of anophelines with human malaria in Thailand.
Trans R Soc Trop Med Hyg 63: 613-619.
Santoro F, Cochrane AH, Nussenzweig V, Nardin EH, Nussenzweig
R, Gwadz RW, Ferreira A 1983. Structural similarities among
the protective antigens of spoorzoites from different species
oof malaria parasites. J Biol Chem 258: 3341-
3345.
Sattabongkot J, Manaeechai N, Rosenberg R 1991. Plasmodium
vivax: gametocyte infectivity of naturally infected Thai
adults. Parasitology 102: 27-31.
Schall JJ 1989. The sex ratio of Plasmodium
gametocytes. Parasitology 98: 343-350.
Schapira A, Beales PF, Halloran ME 1993. Malaria: living with
drug resistance. Parasitol Today 9: 168-174.
Scherf A, Behr C, Sarthou JL, Pla M, Rogier C, Trape JF,
Pereira da Silva L, Dubois P 1993. Immune response in mouse
and malaria-exposed humans to peptides derived from Pf11-1, a
highly repetitive megadalton protein of Plasmodium
falciparum. Eur J Immunol 23: 1574-1581.
Scherf A, Petersen C, Carter R, Alano P, Nellson R, Aikawa M,
Mattei D, Pereira da Silava, L. 1992. Characterization of a
Plasmodium falciparum mutant that has deleted the
majority of the gametocyte-specific Pf11-1 locus. Mem Inst
Oswaldo Cruz 87(Suppl. III): 91-94.
Schneweis S, Maier WA, Seitz HM 1991. Haemolysis of infected
erythrocytes - A trigger for formation of Plasmodium
falciparum gametocytes. Parasitol Res 77:
458-460.
Schulman S, Oppenheim JD, Vanderberg JP 1980. Plasmodium
berghei and Plasmodium knowlesi: serum binding to
sporozoites. Exp Parasitol 49: 420-429.
Seitz HM, Maier WA, Rottok M, Becker-Feldman H 1987.
Concomittant infections of Anopheles stephensi with
Plasmodium berghei and Serratia marcesens
additive detrimental effects. Z Bakteriol Mikrobiol
Hyg 266: 155-166.
Shahabuddin M, Toyoshima T, Aikawa M, Kaslow DC 1993.
Transmission-blocking acitivity of a chitinase inhibitor
activation of malarial parasite chitinase by mosquito
protease. Proc Natl Acad Sci USA 90: 4266-
4270.
Shute PG 1945. An investigation into the number of sporozoites
found in the salivary glands of Anopheles mosquitoes.
Trans R Soc Trop Med Hyg 38: 493-498.
Shute PG, Marion M 1951. A study of gametocytes in a West
Africa strain of Plamodium falciparum. Trans R Soc
Trop Med Hyg 44: 421-438.
Shute PG, Maryon M 1954. The effect of pyrimethamine
(Daraprim) on the gametocytes and oocysts of Plasmodium
falciparum and Plasmodium vivax. Trans R Soc
Trop Med Hyg 48: 50-63.
Sieber KP, Huber M, Kaslow D, Banks SM, Torii M, Aikawa M,
Miller LH 1991. The peritrophic membrane as a barrier: its
penetration by Plasmodium gallinaceum and the effect of
a monoclonal antibody to ookinetes. Exp Parasitol
72: 145-156.
Simonetti AB, Billingsley PF, Winger L, Sinden RE 1993.
Kinetics of expression of two major Plasmodium berghei
antigens in the msquito vector Anopheles stephensi.
J Euk Microbiol (J Protozool) 40: 569-576.
Sinden RE 1975. The sporogonic cycle of Plasmodium yoelii
nigeriensis: a scanning electron microscope study.
Protistologica 11: 31-39.
Sinden RE 1983. The cell biology of sexual development in
Plasmodium. Parasitology 86: 7-28.
Sinden RE 1984. The biology of Plasmodium in the
mosquito. Experientia 40: 1330-1343.
Sinden RE 1985. A cell biologist's view of host cell
recognition and invasion by malarial parasites. Trans R
Soc Trop Med Hyg 79: 598-605.
Sinden RE 1978. Cell Biology, p. 85-186. In R Killick-Kendrick
& WA Peters (eds) Rodent Malaria, New York: Academic
Press.
Sinden RE 1989. Recent advances in the parasitology of
malaria. Trans R Soc Trop Med Hyg 83 Suppl:
3-9.
Sinden RE 1991. Asexual blood stages of malaria modulate
gametocyte infectivity to the mosquito vector - possible
implications for control strategies. Parasitology
103: 191-196.
Sinden RE, Butcher GA, Billker O, Fleck SL 1996. Regulation of
infectivity of Plasmodium to the mosquito vector. Adv
Parasitology 38: 54 - 117.
Sinden RE, Croll NA 1975. Citolology and kinetics of
microgametogenesis and fertilization of Plasmodium yoelii
nigeriensis. Parasitology 70: 53-65.
Sinden RE, Garnham PCC 1973. A comparative study on the
ultrastructure of Plasmodium sporozoites within the
oocyst and salivary glands, with particular reference to the
incidence of the micropore. Trans R Soc Trop Med Hyg
67: 631-637.
Sinden RE, Smalley ME 1976. Gametocytes of Plasmodium
falciparum: phagocytosis by leucocytes in vivo and in
vitro. Trans R Soc Trop Med Hyg 70: 344-345.
Sinden RE, Strong K 1978. An ultrastructural study of the
sporogonic development of Plasmodium falciparum in
Anopheles gambiae. Trans R Soc Trop Med Hyg
27: 477-491.
Sinden RE, Winger LA, Hartley RH, Carter HE, Tirawanchai N,
Davies CS, Sluiters JG 1987. Ookinete antigens of
Plasmodium berghei: a light and electron microscopic
immunogold study of the 21kD determinant recognised by
transmission blocking antibodies. Proc R Soc Lond B
230: 443-458.
Sluiters JF, Visser PE, Van Der Kaay, HJ 1986. The
establishment of Plasmodium berghei in mosquitoes of a
refractory and a susceptible line of Anopheles
atroparvus. Z
Parasitenkd 72: 313-322.
Smalley ME 1977. Plasmodium falciparum gametocytes: the
effect of chloroquine on their development. Trans R Soc
Trop Med Hyg 71: 523-529.
Smalley ME, Brown J, Basset NM 1981. The rate of production of
Plasmodium falciparum gametocytes during natural
infections. Trans R Soc Trop Med Hyg 75: 317-
319.
Snewin VA, Premawansa S, Kapilananda GMG, Ratnayaka L, Udagama
PV, Mattei DM, Khouri E, DelGiudice G, Peiris JSM, Mendis KN,
David PH 1995. Transmission blocking immunity in Plasmodium
vivax malaria: antibodies raised against a peptide block
parasite development in the mosquito vector. J Exp Med
181: 357-362.
Snow RW, Schellenberg JRMA, Peshu N, Forster D, Newton CRJC,
Winstanley PA, Mwangi I, Warn PA, Newbold C, Marsh K 1993.
Periodicity and space-time clustering of severe childhood
malaria on the coast of Kenya. Trans R Soc Trop Med Hyg
87: 386-390.
Speer CA, Rosale-Ronquillo MC, Silverman PH 1974. Scanning
electron microscope observations of Plasmodium berghei
ookinetes in primary mosquito cell cultures. J Invert
Pathol 24: 179-183.
Sterling CR, Aikawa M, Vanderberg JP 1973. The passage of
Plasmodium berghei sporozoites through the salivary
glands of Anopheles stephensi: an electron microscope
study. J Parasitol 59: 593-605.
Stewart MJ, Vanderberg JP 1988. Malaria sporozoites leave
behind trails of circumsporozoite protein during gliding.
J Protozool 35:389-393.
Stewart MJ, Vanderberg JP 1991. Malaria sporozoites release
circumsporozoite protein from their apical end and translocate
it along their surface. J Protozool 38: 411-
421.
Stewart MJ, Vanderberg JP 1992. Electron microscopic analysis
of circumsporozoite protein trail formation by gliding malaria
sporozoites. J Protozool 39: 663-671.
Stohler H 1957. Analyse des infektionsverlaufes von
Plasmodium gallinaceum im darme von Aedes
aegypti. Acta Trop 14: 302-352.
Strome CPA, Beaudoin RL 1974. The surface of the malarial
parasite I. Scanning electron microscopy of the oocyst.
Exp Parasitol 36: 131-142.
Syafruddin A, Arakawa R, Kamimura, Kawamoto F 1991.
Penetration of the mosquito midgut wall by the ookinetes of
Plasmodium yoelii nigeriensis. Parasitol Res
77: 230-236.
Syafruddin A, Arakawa R, Kamimura K, Kawamotoo F 1992.
Development of Plasmodium berghei ookinetes to young
oocysts in vitro. J Protozool 39: 333-338.
Targett GAT 1989. Status of malaria vaccine research. J R
Soc Med 82: 52-56.
Targett GAT 1990. Immunity to sexual stages of human malaria
parasites-Immune modulation during natural infections,
antigenic determinants, and the induction of transmission-
blocking immunity. Scand J Infec Dis 76: 79-88.
Terzakis JA 1971. Transformation of the Plasmodium
cynomolgi oocyst. J Protozool 18: 62-73.
Tirawanchai N, Winger LA, Nicholas J, Ssinden RE 1991.
Analysis of immunity induced by the affinity-purified 21-
Kilodalton zygote-ookinete surface antigen of Plasmodium
berghei. Infec Immun 59: 36-44.
Titus RG, Ribeiro JMC 1990. The role of vector saliva in
transmission of arthropod-borne disease. Parasitol
Today 6: 157-160.
Torii M, Nakamura K, Sieber KP, Miller LH, Aikawa M 1992a.
Penetration of the mosquito (Aedes aegypti) midgut wall
by the ookinetes of Plasmodium gallinaceum. J
Protozool 39: 449-454.
Torii M, Yanagi T, Tsuboi T, Kanbara H 1992b. Appearence and
localization of Plasmodium yoelii circumsporozoite
protein during sporogony in Anopheles stephensi. J
Protozool Res 2: 134-140.
Touray MG, Warburg A, Laughinghouse A, Krettli AU, Miller LH
1992. Developmentally regulated infectivity of malaria
sporozoites for mosquito salivary glands and the vertebrate
host. J Exp Med 175: 1607-1612.
Tsuji M, Corradin G, Zavala F 1992. Monoclonal antibodies
recognize a processing dependent epitope present in the mature
CS protein of various plasmodial species. Parasite
Immunol 14: 457-469.
Turner DP, Gregson NA 1982. The cell surface of Plasmodium
gallinaceum sporozoites: microelectrophoretic and lectin-
binding studies. Parasitology 84: 227-238.
Van Houten J 1988. Chemoresponse mechanism toward a molecular
level. J Protozool 35: 241-245.
Vanderberg JP 1974. Studies on the motility of
Plasmodium sporozoites. J Protozool 21:
527-537.
Vanderberg JP 1975. Development of infectivity of the
Plasmodium berghei sporozoite. J Parasitol
61: 43-50.
Vanderberg JP 1977. Plasmodium berghei: quantitation of
sporozoites injected by mosquitoes feeding on a rodent host.
Exp Parasitol 42: 169-181.
Vanderberg JP, Rhodin J, Yoeli M 1967. Electron microscopic
and histochemical studies of sporozoite formation in
Plasmodium berghei. J Protozool 14: 82-
103.
Vaughan JA, Do Rosario V, Leland P, Light J, Woolett R,
Hollingdale MR, Azad AF 1988. Plasmodium falciparum:
ingested anti-sporozoite antibodies affect sporogony in
Anopheles stephensi mosquitoes. Exp Parasitol
66: 171-182.
Vaughan JA, Noden BH, Beier JC 1992. Population dynamics of
Plasmodium falciparum sporogony in laboratory-infected
Anopheles gambiae. J Parasitol 78: 716-724.
Vaughan JA, Noden BH, Beier JC 1994. Sporogonic development of
cultured Plasmodium falciparum in six species of
laboratory-reared Anopheles mosquitoes. Am J Trop
Med Hyg 51: 233-243.
Vermeulen AN, Deursen JV, Brakenhof RH, Lensen THW, Ponnudurai
T, Meuwissen JHET 1986. Characterization of Plasmodium
falciparum sexual stage antigens and their biosynthesis in
synchronized gametocyte cultures. Mol Biochem
Parasitol 20: 155-163.
Vermeulen AN, Ponnudurai T, Beckers PJA, Verhave JP, Smits MA,
Meuwissen JHET 1985a. Sequential expression of antigens on
sexual stages of Plasmodium falciparum accessible to
transmission blocking antibodies in the mosquito. J Exp
Med 162: 1460-1476.
Vermeulen AN, Roeffen WFG, Hendrik JBJ, Ponnudurai T, Beckers
PJA, Meuwissen JHET 1985b. Plasmodium falciparum
transmission blocking antibodies recognize monovalently
expressed epitopes. Develop Biol Standard 62: 91-97.
Vernick KD, Collins H, Gwadz RW 1989a. A general system of
resistance to malaria infection in Anopheles gambiae
controlled by two main genetic loci. Am J Trop Med Hyg
40: 585-592.
Vernick KD, Collins FH 1989b. Association of a
Plasmodium-refractory phenotype with an esterase locus
in Anopheles gambiae. Am J Trop Med Hyg
40: 593-597.
Vickerman K, Cox FEG 1972. Introductory studies in Biology. In
SW Hurry & J Murray (eds), The Protozoa, Albemarie
Street, London.
Warburg A, Miller LH 1992. Sporogonic development of a malaria
parasite in vitro. Science 255: 448-
450.
Warburg A, Schneider I 1993. In vitro culture of the mosquito
stages of Plasmodium falciparum. Exp Parasitol
76: 121-126.
Warburg A, Touray M, Krettli AU, Miller LH 1992. Plasmodium
gallinaceum: antibodies to circumsporozoite protein
prevent sporozoites from invading the salivary glands of
Aedes aegypti. Exp Parasitol 75: 303-
307.
Weathersby AB 1952. The role of the stomach wall in the
exogenous development of Plasmodium gallinaceum as
studied by means of haemocoel injections of susceptible and
refractory mosquitoes. J Infec Dis 91: 198-
205.
Weathersby AB 1954. The ectopic development of malarial
oocysts. Exp Parasitol 3: 538-543.
Weathersby AB 1960. Experimental infection of Aedes
aegypti with exoerythrocytic stages of Plasmodium
gallinaceum. Exp Parasitol 9: 334-337.
Wernsdorfer WH 1991. The development and spread of drug-
resistant malaria. Parasitol Today 7: 297-
303.
Wilkinson RN, Colwell EJ, Neoypatimanond S 1973. Effect of
sulphamethoxazole-trimethoprim on the viability of
Plasmodium falciparum gametocytes. Trans R Soc Trop
Med Hyg 67: 148-149.
Wilkinson, RN, Neypatimanond, S, Gould DJ 1976. Infectivity of
falciparum malaria patients for anopheline mosquitoes befoore
and after chloroquine treatment. Trans R Soc Trop Med
Hyg 70: 306-307.
Winger LA, Tirawanchai N, Nicholas J, Carter HE, Smith JE,
Sinden RE 1988. Ookinete antigens of Plasmodium
berghei. Appearance on the zygote of a Mr 21 kD
determinant identified by transmission blocking monoclonal
antibodies. Parasite Immunol 10: 193-207.
Wirtz RA, Burkot TR, Graves PM, Andre RG 1987. Field
evaluation of enzyme-linked immunosorbent assays for
Plasmodium falciparum and Plasmodium vivax
sporozoites in mosquitoes (Diptera: Culicidae) from Papua New
Guinea. J Med Entomol 24: 433-437.
Yoshida N, Potonjak P, Nussenzweig V, Nussenzweig RS 1981.
Biosynthesis of Pb44, the protective antigen of sporozoites of
Plasmodium berghei. J Exp Med 154:
1225-1236.
Young MD, Burgess RW 1961. The infectivity to mosquitoes of
Plasmodium malariae. Am J Hyg 73: 182-
192.
|
| |||||||||

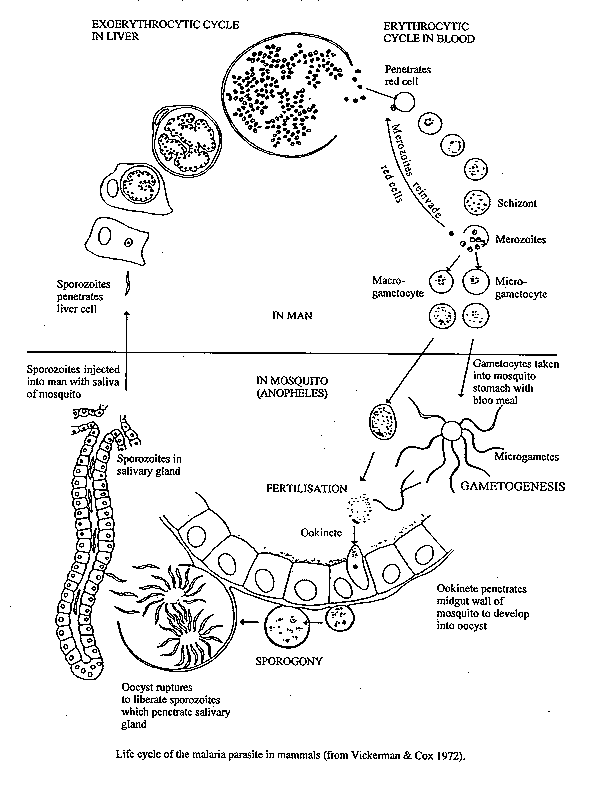{kind=link}