 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Histopathological Study of Experimental and Natural Infections by Trypanosoma cruzi in Didelphis marsupialis
Jo o Carlos Araujo Carreira/^+, Ana Maria Jansen, Maria P Deane^U, Henrique Leonel Lenzi*
Laboratorio de Biologia de Tripanosomatideos, Departamento de
Protozoologia ^*Departamento de Patologia, Instituto Oswaldo
Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ,
Brasil
Received 30 November 1995
Code Number: OC96110
Size of Files:
Text: 35.1K
Graphics: photographs (jpg) - 361.9K[TABLES AND FIGURES AT END OF TEXT]
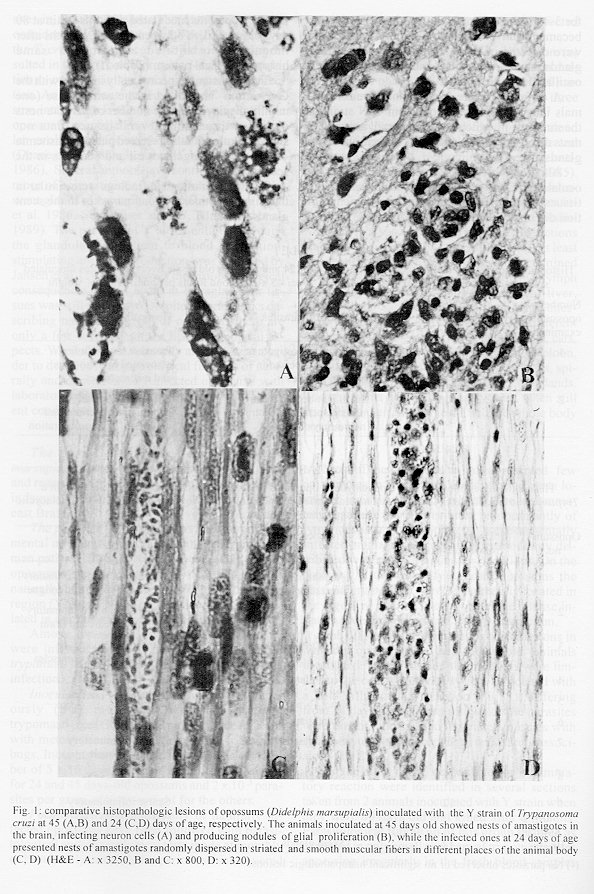
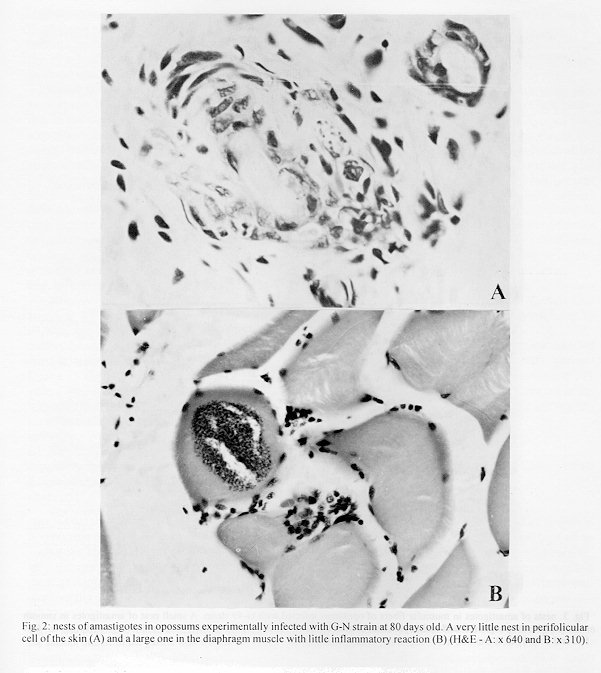
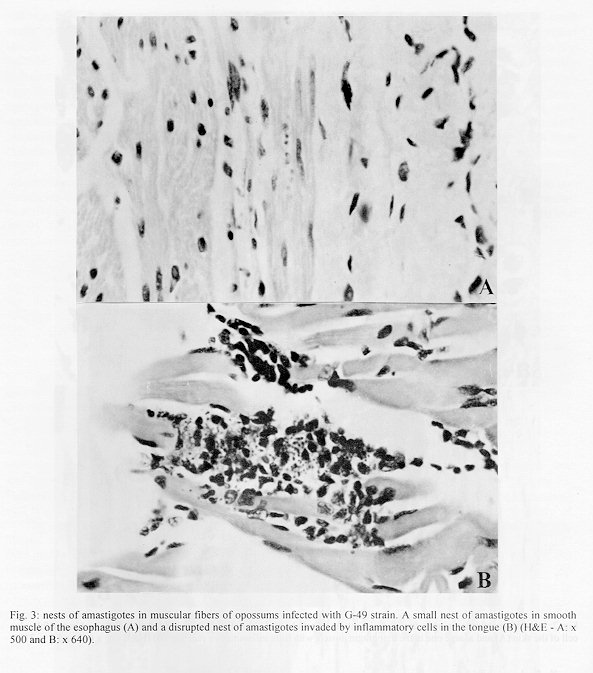
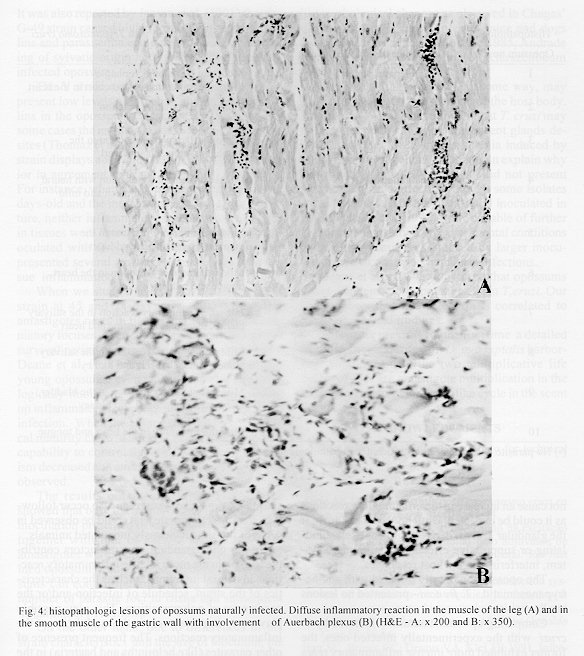
Didelphis marsupialis, the most important sylvatic reservoir of Trypanosoma cruzi, can also maintain in their anal scent glands the multiplicative forms only described in the intestinal tract of triatomine bugs. A study of 21 experimentally and 10 naturally infected opossums with T. cruzi was undertaken in order to establish the histopathological pattern under different conditions. Our results showed that the inflammation was predominantly lymphomacrophagic and more severe in the naturally infected animals but never as intense as those described in Chagas' disease or in other animal models. The parasitism in both groups was always mild with very scarce amastigote nests in the tissues. In the experimentally infected animals, the inflammation was directly related to the presence of amastigotes nests. Four 24 days-old animals, still in embryonic stage, showed multiple amastigotes nests and moderate inflammatory reactions, but even so they survived longer and presented less severe lesions than experimentally infected adult mice. Parasites were found in smooth, cardiac and/or predominantly striated muscles, as well as in nerve cells. Differing from the experimentally infected opossums parasitism in the naturally infected animals predominated in the heart, esophagus and stomach. Parasitism of the scent glands did not affect the histopathological pattern observed in extraglandular tissues. Key words: Trypanosoma cruzi - Didelphis marsupialis - opossum - histopathology Marsupials of the genus Didelphis are considered the most important natural reservoir of Trypanosoma cruzi. The first case of natural infection by T. cruzi in marsupials was reported by Robertson (1929) in D. marsupialis. Further studies have confirmed the importance of this reservoir showing levels of natural infection ranging from 17 to 100% (Rodrigues & Mello 1942, Guimar es & Jansen 1943, McKeever & Gorman 1958, Herrera & Urdaneta-Morales 1992, Travi et al. 1994). Studying natural infections, Zeledon et al. (l970) found only 19 infected out of 100 opossums, detecting few but typical amastigotes in sections of the heart, esophagus, small and large intestine and skeletal muscle. Petana (1969) observed only very few and small amastigote nests in the heart muscle of one naturally infected opossum. Brito and Deane (1966) examined 8 opossums, and although only 1 had amastigotes in the stomach, they observed interstitial inflammatory reactions in the heart, stomach and intestine in all of them. Barr et al. (1991) examining 48 opossums, observed myocarditis in 22, although only 6 exhibited amastigotes nests in the heart and in the tongue. These authors did not mention inflammatory reactions related to T. cruzi in other sites in the animal. More recently Herrera and Urdaneta-Morales (1992) observed only few parasites in the heart of 2 naturally infected opossums. Parasitological studies (Deane et al.1984a, Jansen et al. 1985, Jansen et al. 1991) of opossums experimentally infected with T. cruzi showed that since very young, opossums resist to experimental infections with high inocula and that two types of infection could occur: (1) apparently self- cured, where the results of sub-inoculation (in mice), xenodiagnosis and hemoculture became negative within a few weeks after inoculation, and the immunofluorescent antibody test (IFAT) was always positive at low titers and no invasion of the scent glands could be observed; (2) infection where the parasites were often intermittently recovered through xenodiagnosis and/or hemoculture, after many months; IFAT was persistently positive at high titers and most of the animals presented parasites in the scent glands. These differences were considered by Deane et al. (1984a,b) as related to the T. cruzi-strain, since: Y strain inoculations resulted in the first and F or sylvan-isolates inoculations resulted in the second pattern of infection. An important aspect of the T. cruzi-opossum interaction is the cycle of the parasite in the opossum's scent glands where, T. cruzi shows similar evolutional forms as in the triatomid bugs (Deane et al. 1984a, Lenzi et al. 1984, Deane et al. 1986). Several authors have confirmed the occurrence of epimastigotes and trypomastigotes in the scent glands of naturally infected opossums (Naiff et al. 1986, Steindel et al.1987, Fernandes et al. 1989). The passage of T. cruzi antigens through the glandular epithelium to blood circulation, stimulating antibody production, was observed by Jansen et al. (1988), however the histopathological consequences of this event in extraglandular tissues was still unknown. Despite several works describing natural infection of opossums there are only a few focusing on the histopathological aspects. We undertook our study at this point in order to describe histopathological findings of naturally and experimentally infected opossums with laboratory appraised T. cruzi-strains under different conditions. Materials and methods The host - Opossums, identified as D. marsupialis, were: (a) specimens conceived, born and reared in captivity and, (b) specimens trapped in Jaguanum Island, State of Rio de Janeiro, southeast Brazil. The parasite - T. cruzi strains used for experimental infections were: (a) Y, isolated, from a human patient (Silva & Nussenzweig 1953) and (b) opossums strains G-N and G-49, isolated from naturally infected D. marsupialis in the Amazon region (Yoshida 1983) and in Rio de Janeiro (isolated in our laboratory 1985), respectively. Among opossums trapped in the field, 10 were infected with T. cruzi, 3 with T. (Mega-trypanum) freitasi and 3 were free of trypanosome infection. Inoculations - The animals were subcutaneously (SC) inoculated with bloodstream trypomastigotes (BST) from maintenance mice, or with metacyclic trypomastigotes from triatomine bugs. Inocula were adjusted on the basis of number of 5 x 10^3 parasites per gram of body-weight for 24 and 45 days-old opossums and 2 x 10 ^3 parasites per gram of body-weight for the others. Follow-up - Experimentally or naturally infected opossums have been followed with parasitological and/or serological tests made at monthly or bi-monthly intervals for periods which covered more than one year for some naturally infected animals. Parasitological diagnosis - It was based on: (a) direct blood examination of fresh preparation during the acute phase; (b) hemoculture in NNN medium examined through a period of one to three months. Serology was done on blood samples obtained by vein (femoral or caudal) or heart puncture and the sera were tested by immunoflourescent antibody test (IFAT) described by Jansen et al. (1985). Necropsy - The animals were killed by intramuscular injection of Ketalar and chloroform inhalation. Tissues samples were fixed in formalin-Millonig (Carson et al. 1973) and paraffin sections stained with hematoxilin and eosin (H&E). At least one section of the following tissues was examined from all the animals: tongue, tonsils, thyroid, lymph nodes, lung, heart, esophagus, stomach, liver, spleen, diaphragm muscle, small and large intestines, skeletal muscle, bone marrow of different places, pancreas, eyes, brain, cerebellum, skin, ears, gallbladder, bladder, marsupial pouch, epiploon, mesentery, bulbourethral glands, hypophysis, spinal chord, kidney, genital tracts and scent glands. The opossums were killed very young, when still attached to their mother teats, had the whole body processed. Results Most of the examined animals presented few amastigotes in small, rare and dispersed nests localized mainly in skeletal muscle. The inflammatory reactions were constituted predominantly of lymphocytes and macrophages. In experimentally infected animals the parasites were randomly distributed with mild inflammatory reactions. On the contrary, in the naturally infected opossums the nests of amastigotes were preferentially located in the digestive tract and heart with more intense inflammation and occasional tissue destruction. Table I shows the histopathological lesions in newborn opossums: in the 45 days old animals inoculated with Y strain, the parasites were limited to the central nervous system associated with a mild inflammatory reaction (Fig. 1), differing from 24 days old opossums in which the parasites were randomly dispersed in striated muscles with lymphocytes and macrophages only rarely associated (Fig. 2). On the other hand, no parasites or inflammatory reaction were identified in several sections taken from 2 animals inoculated with Y strain when they were 80 days old and killed after 8 months. Parasitological follow up of this group resulted as expected in only small numbers of trypomastigotes appearing irregularly in the fresh blood samples for 3-4 weeks, after which also the hemocultures became negative. No invasion of the scent glands were detected in fresh preparations of the scent glands content. The total immunoglobulins titers oscillated between 1:40-1:80. A different picture could be observed in animals inoculated with the opossum G-N strain at the same age in which we found small amastigote nests distributed in striated muscle fibers and scent glands. Although only one out of the 10 opossums inoculated with G-49 strain presented parasites in the tissues (Fig. 3), the duration of the chronic infection did not determine the severity of the lesions. Indeed, opossums inoculated with this strain at 80 days old and killed between 5 and 12 months after inoculation exhibited almost the same histopathological pattern (Table II). Two opossums experimentally infected with the G-49 strain and killed in the acute phase (one month) displayed a large number of parasite nests dispersed throughout several tissues (data not shown). Both animals presented patent parasitemia - up to 1 x 10^5 flagellates/ml and parasites in the scent glands. The histopathological findings were similar in the animals with or without parasites in the scent glands (Table II). Only 4 out of 10 naturally infected opossums showed amastigotes in the tissues. The heart, stomach and esophagus were involved in 2 of them; bladder and ureter in 1. Inflammatory reaction was focal, without fibrosis and directly related to the intensity of tissue parasitism which was usually light and more concentrated in the heart and digestive tract (Fig. 4, Table III). In the heart there was focal and interstitial myocarditis, mainly in atria involving the subendothelial layer. In a single case there was inflammation in intramural ganglion. In the stomach the infiltrate invaded the Auerbach plexus and muscular layers. A similar inflammatory reaction pattern lower in intensity was observed in the six T. cruzi infected animals with no detectable tissue parasitism (Fig. 4). Besides the natural infection by T. cruzi, all the opossums harbored different species of helmints as well as protozoa, bacteria and fungi. All the naturally infected animals presented subpatent parasitemias, positive hemocultures and no parasite in the scent glands. The serological titers (IFAT) in these animals oscillated between 1:80-1:160. Opossums naturally infected with T. freitasi as well as the uninfected ones displayed some nonspecific lesions - probably due to the presence of other parasites such as helminths and bacteria that were observed in most of animals. Discussion Opossum probably starts to resist to infections with Y strain from the 45th day of life on since all 24 days old inoculated animals died in contrast to the survival of the older ones (Deane et al. 1984c). Although the opossums acquire immunological maturity from the 80th day of age on (Block 1964), younger animals (45 days old), were able to control the infection by T. cruzi (Deane et al. 1984c), suggesting that other mechanisms, probably nonspecific, are involved in the efficient control of the infection as also suggested by Jansen et al. (1991), at least in animals at this age. Indeed, when 2 pouch young opossums (@80 days) were inoculated subcutaneously with 2 x 10^3 blood forms of Y strain per gram of body weight, and killed at 8 month after infection, parasites and tissue lesions were not detected. Only few parasites and a mild inflammatory reaction in the central nervous system was observed in the 45 days old infected opossums necropsed 2 months after the inoculation with Y strain trypomastigote (Table I). The parasitological follow-up correlated entirely with the histopathological findings depending also on the inoculated strain. Opossums experimentally infected with Y strain displayed a short period of patent parasitemia with rare positive fresh blood examinations and no scent glands parasitism. On the contrary, the infections with G-N strain resulted in high parasitemia up to 1x10^7 parasites/ml and heavily parasitized scent glands. The monthly hemocultures done during the subpatent period were consistently positive in the G-N inoculated animals, contrasting to those inoculated with the Y strain (data not shown). It is known that T. cruzi antigens pass through the scent glands wall to the blood (Jansen et al. 1988). However, the fact that several animals with heavily parasitized scent glands did not show or presented only scarse inflammatory reactions in the tissues suggests that these circulating antigens do not cause an increase in the inflammatory reactions as it could be expected. Until now, it is unknown if the glandular T. cruzi derived antigens exert stimulating or suppressive effects on the immune system, interfering on the host responses. The opossums naturally infected with another trypanosomatid - T. freitasi - presented no lesions at all. Comparing opossums naturally infected by T. cruzi with the experimentally infected ones, the former exhibited more intense inflammatory reactions in tissues and a tendency for parasitism to occur in the heart and digestive tract. The pattern of inflammatory reactions in both groups was qualitatively similar, composed of macrophages and lymphocytes with occasional small numbers of eosinophils and/or plasma cells. Parasitism in the digestive system by T. cruzi in natural infections of opossums was already observed (Brito & Deane 1966, Zeledon 1970, Barr et al. 1991). Brito and Deane (1966) suggested that it probably could occur through oral infections. Although this hypothesis can be true, T. cruzi parasites in the digestive system can also occur following other infection route as it could be observed in some of the subcutaneously inoculated animals. In our understanding, several factors contribute to explain the more intense inflammatory reactions in natural infections, such as the characteristics of the strain, schedule of infection and/or the occurrence of polyparasitism. This may possibly play a very important role in the modulation of the inflammatory reactions. The frequent presence of other parasites (like helminths and bacteria) in the organism of the animals could cause a more severe immune response due to superimposition of immunological reactions stimulated by different agents. Favoring this hypothesis is the fact that all the animals, caught in the wild and heavily infected by helminths and other parasites, presented more pronounced histopathologic lesions than those animals born in the laboratory. It was reported that the opossum is able to set up a humoral response to T. cruzi antigens and that the levels of serum specific antibodies is dependent on the strain of parasite (Jansen et al. 1985). It was also reported by Jansen et al. (1991) that the G-49 strain caused higher levels of immunoglobulins and parasitemia compared to G-327, both being of sylvatic origin and isolated from naturally infected opossums.
The Y strain, on the other hand, is known to present low levels of infection and immunoglobulins in the opossums (Jansen et al. 1985), and in some cases the animals rid themselves of the parasites (Thomaz et al. 1984). We observed that the Y strain displays correlated histopathological behavior in agreement with parasitological conditions. For instance, when the opossums were close to 80 days-old and the immune system had become mature, neither inflammatory reactions nor parasites in tissues were detected. Otherwise, opossums inoculated with G-N strain in the same age group presented several amastigotes nests and light tissue inflammation. When we studied animals inoculated with Y strain at 45 and 24 days, we observed several amastigotes nests in tissues and scattered inflammatory focuses. Even so, the opossums had a longer survival period compared to mice, as observed by Deane et al. (1984c). These results indicate that young opossums, even when they were immunologically immature, they were capable of setting up inflammatory responses sufficient to control the infection. When the animal reached immunological maturity close to 80 days of age, they acquired capability to control the infection. Tissue parasitism decreased and amastigotes nests were no longer observed. The results obtained with the G-49 strain showed that only the animal killed 5 months after inoculation presented tissue parasitism. This data, together with that of natural infection (where amastigotes were found only in 4 of the 10 examined opossums), suggest that sometimes the opossums can control the infection by decreasing the number of parasite foci in tissue and subsequent inflammatory reactions. On the other hand, due to differences of the strain characteristics and the size of inoculum, the opossums do not always control the colonization of the tissue by the parasite. The parasites can remain in the tissues for a long time, with very limited inflammatory reactions, as we could observe in animals infected with the G-N strain.
T. cruzi is a very heterogenous species; consequently, the infection by this parasite may be very complex, depending on the host and parasite factors. Our observations on the behavior of T. cruzi in the D. marsupialis indicate that even those naturally infected animals showing more severe inflammatory reactions did not present a pronounced histopathological changes as observed in Chagas' disease, and in T. cruzi infected mice and dogs (Andrade & Carvalho 1969, Aluja 1985, Andrade & Sadigursky 1987, Molina & Kierszenbaum 1988). The size of the inoculum, in some way, may modulate the parasite dispersion in the host body. Jansen et al. (1991), have shown that T. cruzi may or may not reach the opossums' scent glands depending on the levels of parasitemia induced by the size of the inoculum. This fact can explain why our naturally infected opossums did not present anal scent glands infection, though some isolates when obtained from the glands and inoculated in others (Jansen et al.1991), were capable of further glandular colonization. In experimental conditions higher parasitemia was induced by larger inoculum than that observed in natural infections. Deane et al. (1984a), observed that opossums select different sub-populations from T.cruzi. Our study shows that this selection is correlated to histopathological findings. This paper presents for the first time a detailed histopathological study of D. marsupialis harboring simultaneously the two multiplicative life cycles of T. cruzi: amastigote multiplication in the tissues and triatomine vector-like cycle in the scent glands. Acknowledgments To Luzia Fatima Goncalves Caputo for technical assistance.
References Aluja AS 1985. Miocarditis por Trypanosoma cruzi en un perro. Veterinaria Mex 16: 41-44. Andrade SG, Carvalho ML 1969. Efeito da excitac o do sistema reticulo-endotelial pelo adjuvante de Freud, na doenca de Chagas experimental. Rev Inst Med Trop S o Paulo 11: 229-235. Andrade SG, Sadigursky M 1987. The conduction system of the heart in mice chronically infected with Trypanosoma cruzi: histopathological lesions and electrocardiographic correlations. Mem Inst Oswaldo Cruz 82: 59-66. Barr SC, Brown CC, Dennis VA, Klei TR 1991. Infection of inbred mice with 3 Trypanosoma cruzi isolated from Louisiana (USA) mammals. J Parasitol 76: 918-921. Block M 1964. VI The blood forming tissues and the blood of the newborn opossum (Didelphis marsupialis). Ergeb Anat Entwicklungsgesh 37: 237- 366. Brito T, Deane LM 1966. Aspectos anatomo-patologicos da infecc o natural de alguns animais silvestres pelo Trypanosoma cruzi. Rev Inst Med Trop S o Paulo 8: 79-82. Carson FL, James MS, Martin JH, Lynn JA 1973. Formalin fixation for electron microscopy: a re-evaluation. Am J Clin Path 59: 365-373. Deane MP, Jansen AM, Lenzi HL 1984a. Trypanosoma cruzi: vertebrate and invertebrate cycles in the same mammal host, the opossum Didelphis marsupialis. Mem Inst Oswaldo Cruz 79: 513-515. Deane MP, Jansen AM, Mangia RHR, Goncalves AM, Morel CM 1984b. Are our laboratory strain representative samples of Trypanosoma cruzi populations that circulate in nature? Mem Inst Oswaldo Cruz (Suppl) 79: 19- 24. Deane MP, Jansen AM, Mangia RHR, Lenzi HL 1984c. A study of experimental infection of the opossum Didelphis marsupialis. Annals of the XI Annual Meeting of Basic Research on Chagas' Disease (Abstract BI-39). Deane MP, Lenzi HL, Jansen AM 1986. Double development cycle of Trypanosoma cruzi in the opossum. Parasitol Today 2: 146-147. Fernandes AJ, Diotaiuti L, Pinto Dias JC, Romanha AJ, Chiari E 1989. Infecc o natural das glandulas anais de gambas Didelphis marsupialis pelo Trypanosoma cruzi no municipio de Bambui - MG. Mem Inst Oswaldo Cruz 84: 87- 93. Guimar es FN, Jansen G 1943. Novo transmissor silvestre do Trypanosoma (Schizotrypanum) cruzi (Chagas 1909). Mem Inst Oswaldo Cruz 38: 473-441. Herrera L, Urdaneta-Morales S 1992. Didelphis marsupialis: a primary reservoir of Trypanosoma cruzi in urban areas of Caracas, Venezuela. Ann Trop Med Parasitol 86: 607-612. Jansen AM, Carreira JCA, Deane MP 1988. Infection of a mammal by monogenetic insect trypanosomatids (Kinetoplastida,Trypanosomatidae). Mem Inst Oswaldo Cruz 83: 271-272. Jansen AM, Galv o BC, Moriearty PL, Deane MP 1985. Trypanosoma cruzi in Didelphis marsupialis: an indirect fluorescent antibory test for diagnosis and follow up of natural and experimental infections. Trans R Soc Trop Med Hyg 79: 474-476. Jansen AM, Leon L, Machado GM, Da Silva MH, Souza-Le o S, Deane MP 1991 Trypanosoma cruzi in the opossum Didelphis marsupialis: Parasitological and serological follow-up of the acute infection. Exp Parasitol 73: 249-259. Lenzi HL, Jansen AM, Deane MP 1984.The recent discovery of what might be a primordial escape mechanism for Trypanosoma cruzi. Mem Inst Oswaldo Cruz (Suppl) 79: 7- 12. McKeever J, Gorman L 1958. Occurrence of a Trypanosoma cruzi-like organism in some mammals from Southwestern Georgia and Northwestern Florida. J Parasitol 44: 583- 587. Molina HA, Kierszenbaum F 1988. Kinetics of development of inflammatory lesions in myocardial and skeletal muscle in experimental Trypanosoma cruzi infection. J Parasitol 74: 370-374. Naiff RD, Naiff MF, Barret TV, Arias JR 1986. Trypanosoma cruzi nas glandulas anais do Didelphis marsupialis: primeiro registro de infecc es naturais. Annals of the X Meeting of the Brazilian Society of Parasitology (Abstract - 165) . Petana WB 1969. American trypanosomiasis in British Honduras - VI - A natural infection with Trypanosoma (Schizotrypanum) cruzi in the opossum Didelphis marsupialis (Marsupialia, Didelphoidea) and experimental investigations of different wild-animal species as possible reservoirs for the parasite. Ann Trop Med Parasitol 63: 47-56. Robertson A 1929. Note on trypanosome morphologically similar to Trypanosoma cruzi Chagas, 1909, found in opossum, Didelphis marsupialis captured at Tela, Honduras, Central America. XVIII^ th Ann Rep Med Dept Co. 293 - 310. Rodrigues BA, Mello GB 1942. Contribuic o ao estudo da tripanosomiase americana. Mem Inst Oswaldo Cruz 37: 77- 94. Silva LIP, Nussenzweig W 1953. Sobre uma cepa de Trypanosoma cruzi altamente virulenta para camundongo branco. Folia Clin Biol 20: 191-208. Steindel M, Scholz AF, Toma HK, Schlemper Jr BR 1987. Parasitism of blood and anal glands of naturally infected opossum (Didelphis marsupialis) from the Arvoredo Island Santa Catarina by Trypanosoma cruzi. Mem Inst Oswaldo Cruz (Suppl) 82: 66. Thomaz N, Jansen AM, Deane MP 1984. Trypanosoma cruzi. The complement-mediated lysis (Coml) in experimentally infected opossum Didelphis marsupialis. Annals of the XI Annual Meeting of Basic Research on Chagas' Disease (Abstract, I-48), p.116. Travi BL, Jaramilllo C, Montoya J, Segura I, Zea A, Goncalves A, Velez ID 1994. Didelphis marsupialis, an important reservoir of Trypanosoma (Schizotrypanum) cruzi and Leishmania (Leishmania) chagasi in Colombia. Am J Trop Med Hyg 50: 557-565. Yoshida N 1983. Surface antigens of metacyclic trypomastigotes of Trypanosoma cruzi. Infect Immun 40: 836- 839. Zeledon R, Solono G, Saenzs G, Swatzwelder JC 1970. Wild reservoir of Trypanosoma (Schizotrypanum) cruzi with special mention of the opossum, Didelphis marsupialis, and its role in the epidemiology of Chagas' disease in an epidemic area of Costa Rica. J Parasitol 56: 38. --------------------------------------------------------------
Fig. 2: nests of amastigotes in opossums experimentally infected with G-N strain at 80 days old. A very little nest in perifolicular cell of the skin (A) and a large one in the diaphragm muscle with little inflammatory reaction (B) (H&E - A: x 640 and B: x 310). Fig. 3: nests of amastigotes in muscular fibers of opossums infected with G-49 strain. A small nest of amastigotes in smooth muscle of the esophagus (A) and a disrupted nest of amastigotes invaded by inflammatory cells in the tongue (B) (H&E - A: x 500 and B: x 640). Fig. 4: histopathologic lesions of opossums naturally infected. Diffuse inflammatory reaction in the muscle of the leg (A) and in the smooth muscle of the gastric wall with involvement of Auerbach plexus (B) (H&E - A: x 200 and B: x 350).
------------------------------------------------------------- Copyright 1996 Fundacao Oswaldo Cruz
The following images related to this document are available:Photo images[oc96110d.jpg] [oc96110a.jpg] [oc96110c.jpg] [oc96110b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}