 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Antileishmanial Action of Organometallic Complexes of Pt(II) and Rh(I) CM Mesa-Valle/+, V Moraleda-Lindez, D Craciunescu*, A Osuna**
Departamento de Biologia Aplicada, Facultad de Ciencias
Experimentales, Universidad de Almeria, 04120 Almeria,
Espana
Received 26 September 1995
Code Number: OC96113
Size of Files:
Text: 33.0K
Graphics: line drawings (gif) - 19.0K
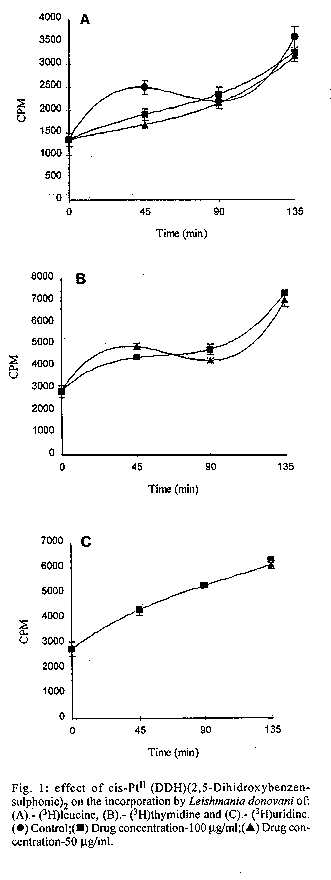
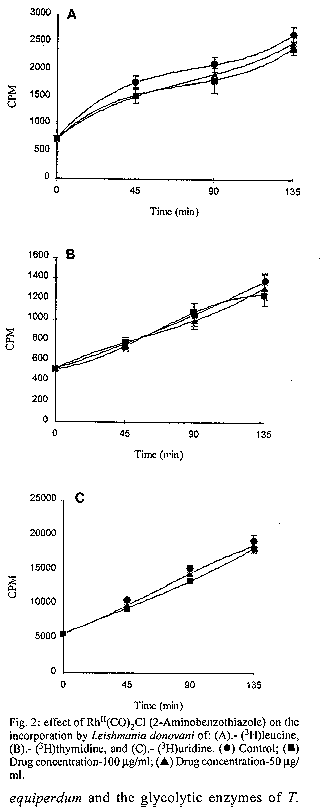
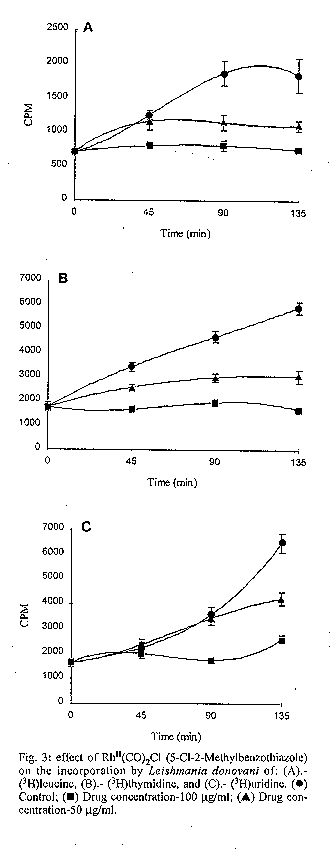
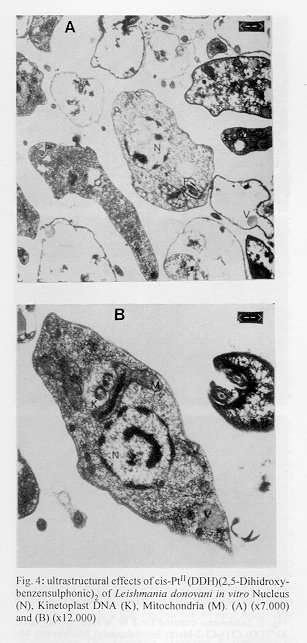
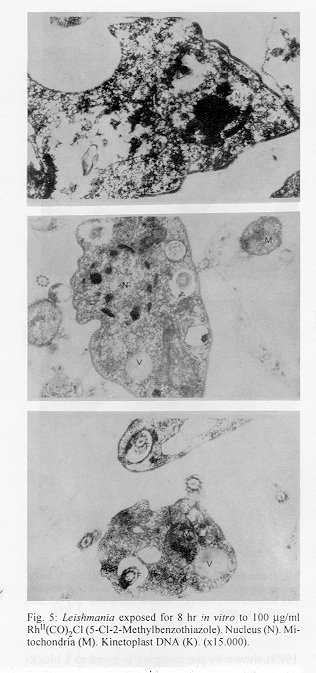
photographs (jpg) - 156.7K[TABLES AND FIGURES AT END OF TEXT] The three organometallic complexes [(Cis-Pt^II (DDH) (2,5- Dihidroxibenzensulfonic)^2, Rh^I (CO)2 Cl(2- Aminobenzothiazole) and Rh^I (CO)2 Cl(5-Cl-2- Methilbenzothiazole)] used in this study had been previously found to have a high in vitro activity against promastigote and amastigote like forms of Leishmania donovani. Here, the cytotoxic effect of these new organometallic complexes on the J-774 macrophages were studied. Only the Rh^I(CO)2 Cl (2-Aminobenzothiazole) complex induced substantial toxicity in the cells. Also, we assayed the effect of this complex on the parasite s biosynthesis of macromolecules. The Rh^I(CO)2Cl (5-Cl-2- Methylbenzothiazole) complex inhibited DNA, RNA, and protein synthesis. On the other hand, the two other compounds tested did not inhibit the incorporation of radioactive precursors. Finally important ultrastructural alterations in the parasites treated with the two non-cytotoxic complexes were observed. Key words: Organometallic complexes - Leishmania donovani - in vitro toxicity - mechanism of action Leishmaniasis is a parasitic disease which affects 12 millon people worldwide (Modabber 1987) and is produced by a species of the genus Leishmania, a protozoo belonging to the family Trypanosomatidae. Manifestations of the illness include more or less generalized cutaneous lesions, or, in the case of visceral leishmaniasis (kala-azar), caused by the species L. donovani or L. infantum, phagocytes of internal organs and tissues (spleen, liver, bone marrow, etc.) are attacked. The drugs for treating these parasitoses have traditionally been pentavalent antimonials, aromatic diamidines and fungicides such as amphotericine B. However, these are extremely toxic and cause a great number of side effects (WHO 1984). Many recent efforts have been made to synthesise and evaluate alternative compounds for treating these parasites (Berman 1988, Croft 1988, Bacchi et al. 1991, Marr 1991). In the last few years, certain metal complexes have proven anti-tumoral against such protozoan parasites as Trypanosoma cruzi, T. rhodesiense and L. donovani. One property that the tumor cells share with the trypanosomatids is rapid multiplication (Kinnamon et al. 1979, Englund et al. 1982, Wysor et al. 1982, Farrell et al. 1984). The anti-trypanosomal activity of a complex with known anti- cancer activity, cis-platino (neoplatin) has been studied by Balber et al. (1985). Other metal complexes, studied by Farrell et al. (1984), Craciunescu et al. (1988a, b, 1990 a, b), Zinststag et al. (1991), Croft et al. (1992), Sanchez- Delgado et al. (1993), and others, have shown potential anti- cancer activity with metals Pt(II), Pt(IV), Ru(II), Ru(III), Rh(I), Rh(III), Ir(I), Ir(II), Os(II) and Os(III) Ir(IV), Pt(II) and Rh(I). In the present work, we have selected three organometallic complexes which have previously shown in vitro activity against the promastigote forms of L. donovani and have also shown a similar activity against the amastigote like forms, these being Pt(II): Cis-Pt(DDH)(Ac. 2,5-Dihydroxi- benzensulphonic)^2 and those of Rh(I): Rh(CO)2 Cl (5Cl-2- Methylbenzothiazole), Rh(CO)^ 2 Cl (2-Aminobenzothiazole) (Mesa-Valle et al. 1989, 1993). First, we assayed the in vitro toxicity of the complexes for the cells of the strain J-774, before evaluating the effect exerted on the parasite s biosynthesis of macromolecules. A study was made of the ultrastructural alterations caused by the two complexes which did not induce toxicity on the cell line assayed. Materials and methods The parasite - The strain of L. donovani used was LCR-L 133 (Leishmania Reference Center, Jerusalem), isolated in 1967 from a human case of kala-azar in Begemder (Ethiopia) and maintened in our laboratory until 1980 by culture in Modified NNN Medium (Mesa-Valle et al. 1993).
The organometallic complexes - The complexes assayed are square, flat organometal compounds: cis-Pt(II) (DDH) (Ac. 2,5-Dihidroxybenzensul-fonic)^2 where DDH is 1,2 diaminocyclohexane; Rh(I)(CO)2 Cl (2- Aminobenzothiazole) and Rh(I)(CO)2 Cl (5-Cl-2- Methylbenzothiazole). The complexes were synthesised according to the methods described by Craciunescu et al. (1985, 1986). Parasite cultures - The promastigote forms of L. donovani were cultured in TC 199 medium (Gibco) supplemented with 30% foetal bovine serum (Gibco) previously inactivated at 56 C for 30 min (SBFI). The amastigote like forms were obtained in vitro by the methods described previously by Castilla et al. (1995). The viability of the forms were determined by fluorescence microscopy after staining the organism with acridine orange 15mg/ml during 15 min at 38 C. For the assays, we used parasites in the exponential phase of growth. The organisms were counted in a Neubauer haemocytometric chamber and the number was adjusted to 1 x 10^6 /ml. Cytotoxicity assay - The macrophage line used, J-774, was cultivated in RPMI medium. When the macrophages had formed a monolayer, the cells were labelled with Cr^51 (50 mCi/ml of final concentration), following the technique of Fulfor et al. (1986). Next, the cells were treated with the different complexes at 100, 10 and 1 mg/ml for 6, 12 and 24 hr. The culture medium was withdrawn and transferred to Eppendorf tubes; SDS-NaOH was added to adhering cells, and this was transferred with the cell remains to new Eppendorf tubes. Finally, in a Beckman g scintillation counter, the gamma radiation in the supernatant and in the pellets was measured, and the percentage of specific release of Cr^51 was calculated using the following values:
CPMSCont
PSL = --------------------- x 100
CPMSCont + CPMPCont
CPMSPro
PLS = ----------------------- x 100
CPMSPro + CPMPPro
PSL= percentage of spontaneous liberation; PSL= percentage of
specific liberation; CPM SCont= number of counts per minute of
the supernatant control; CPM PCpn= number of counts per minute
of pellet control; CPM Spro= number of counts per minute of
the supernatant problem; CPM PPro= number of counts per minute
of pellet problem.
The effect of the metal complexes on the incorporation of [^3H] thymidine, uridine and leucine in the flagellate forms of the parasite - The effect of the organometallic complexes on parasite biosynthesis was measured following the technique described by part of our research group (Gonzalez et al. 1989). The complexes were assayed at concentrations of 100 and 50 mg/ml and were added to the cultures of the Leishmania promastigote forms. The precursors used were thymidine (6-^3H, specific activity 20-30 Ci/mmol), uridine (5-^3H, specific activity Ci/mmol) and leucine (4.5-^3H, specific activity 45- 70 Ci/mmol). A concentration of 5 mCi/ml was added to the culture medium. The recovery of the labelled products, and indirectly the biosynthesis of the macromolecules, were measured after the precipitation of the macromolecules with TCA 10% and filtration in a vacuum through GFC (Watman) filters at 0, 45, 90 and 135 min. The incorporation of the complexes per measure of radioactivity was carried out in a Beckman b Spectrometer using PPO PPOP Cotell. Electron-microscope transmission - To examine the alterations in parasites caused by the metal- drug complexes we treated the parasites in culture with the complexes at a concentration of 100 mg/ml for 8 hr at 28 C. Afterwards, the samples were centrifuged and the pellet was fixed in glutaraldehyde at 2.5% in cacodylate buffer at pH 7.2 and examined under the electron microscope (TEM) following the technique of Osuna et al. (1983). Results Table 1 shows the effect in the parasite forms after treatment with the three compounds. Table II shows the results obtained on treating the macrophage cell line J-774 with the three organometal complexes. Only the complex Rh(I), with the ligand 2-Aminobenzothiazole, induced substantial toxicity to the cells, reflected in 24.2% specific liberation of Cr^51 at 24 hr of treatment and at a concentration of 100 mg/ml. It is noteworthy that this complex showed greater activity in vitro against the parasites, the percentage of inhibition of growth being 97% at 72 hr and 100 mg/ml (Mesa-Valle et al. 1993). Fig. 1 indicates the effects produced by the complex cis-Pt(II)(DDH)(2,5-dihydroxybenzen-sulfonic)^2 on the incorporation of [^3H] thymidine, [^3H] leucine and [^3H] uridine, showing that this complex does not inhibit DNA, RNA or protein synthesis. Similar results were obtained for the complex Rh(I): Rh(CO)2 Cl (2-Aminobenzothia-zole) (Fig. 2), while the complex Rh(CO)2 Cl (5-Cl-2- Methylbenzothiazole) acts on all three levels, decreasing the incorporation of the three radioactive precursors at the two concentrations used, 50 and 100 mg/ml. Thus, at the greater concentration, the inhibtion in thymidine incorporation was 72% at 135 min, while that of leucine and uridine was 60% (Fig. 3). The ultrastructural alterations at the nuclear chromatin level in L. donovani caused by the Pt(II) complex are shown in Fig. 4. Here, a discontinuity of the double nuclear membrane seems to appear. There is a great quantity of lipid vacuoles, as well as the formation of myeline configurations in tissues both at the cytoplasm and the nuclear levels, all showing a high degree of cellular degeneration. Fig 5 and Fig 6 show the effect at the ultrastructural level of the complex Rh(I)(CO)2 Cl (5-Cl-2-Methylbenzotiazole), with similar effects appreciable for complex Pt(II). Discussion Metal complexes with anti-tumor action, such as cis-Pt (neoplatin) have demonstrated their effectiveness in the treatment of trypanosomiasis (Kinnamon et al. 1979, Wysor et al. 1982). Analogues of this complex have been tested against different species of Trypanosoma (Farrell et al. 1984, Ruiz-Perez et al. 1986, 1987) and against Leishmania (Mesa-Valle et al. 1989, 1993). Of the complexes assayed in the present work, the Pt(II) complex Pt(DDH)(2,5-Dihydroxy-benzensulfonic)^2 is a structural analogue of the first (neoplatin). In the same way, this complex is obtained by replacing the NH^3 groups of the neoplatin with (DDH), and the Cl with a ligand of greater liposolubility, (2,5-Dihydroxybenzensulfonic), joined to Pt by a bidentate bond. This structure increases the dynamic stability of the complex, attributable to its greater anti- neoplast effectiveness on the stereochemical disposition of the complex for the interaction with the DNA. Thus, this complex showed activity against T. brucei and T. rhodesiense (Craciunescu et al. 1986), and some structural analogues showed activity against epimastigote forms of T. cruzi (Ruiz-Perez et al. 1986, 1987) and against promastigote forms of L. donovani (Mesa-Valle et al. 1989, 1991). Willson et al. (1992) showed that derivatives of sulphonic acid inhibit the growth of T. equiperdum and the glycolytic enzymes of T. brucei. Glycolysis is the essential mechanism in obtaining energy in the protozoa of the family Trypanosomatidae and occurs in the specific cytosolic bodies called glycosomes (Opperdoes et al. 1984). The three organometallic complexes showed an in vitro antiproliferative activity against both the flagellates and amastigote like forms (Table I). The highest efectivity was obtained at the highest concentration and longest time tested. In an attempt to ascertain whether the leishmanicidal activity in vitro (Mesa-Valle et al. 1989) shown by the complex is owed to a blockage in the biosynthesis of macromolecules, we studied the levels of incorporation of [^3H] thymidine, [^3H] leucine and [^3H] uridine. However, the data obtained with this complex do not coincide with those observed previously by us for other complexes with analogous structures used against T. cruzi (Ruiz-Perez et al. 1987). In the promastigote forms of L. donovani, this complex does not significantly inhibit the incorporation of the labelled analogues by the parasite. At the ultrastructural level, treatment of the promastigote forms for 8 hr with the complex produces abnormal condensations of the chromatin and intranuclear vacuoles with membrane alterations, giving the appearance of mitochondrial fingerprint structures. The mitochondria appear swollen and mitochondrial crests are highly evident in the matrix. No alterations appear at the plasma-membrane level. The complexes Rh, Rh(I) and Rh(III) have shown anti-tumor and anti-bacterial activity similar to that of the derivatives cis-Pt(II) (Dehand & Jordanov 1976, Craciunescu et al. 1986, 1987, 1990a, 1990b). Some of the complexes with greater anti- cancer activity have been found to be strong inhibitors of Ornitine Decarboxylase (Sjoerdsma & Schechter 1984), a target enzyme with a great number of active complexes against various species of Trypanosoma and Leishmania (Bacchi et al. 1980, Fouce et al. 1991). In addition, the ligands used in our case were derived from benzothiazole, a structure in which some of its derivatives have shown promising anti-microbial anti-fungus and anti- parasite activity (Hisano et al. 1982, Haugwitz et al. 1982, Zahner et al. 1991a, b, Yalcin & Sene, 1993, Bujdakova et al. 1993), especially as inhibitors of the synthesis of ergosterol (Urbina et al. 1991), the major sterol in the membranes of fungi, yeasts and trypanosomatids. Known inhibitors of ergosterol synthesis, such as ketoconazol (Goad et al. 1989) show intense anti-leishmania activity (Goad et al. 1985). The complexes from Rh(I) tested by us have a flat, square configuration similar to the structure of neoplatin. Ruiz- Perez et al. (1987) studied the action of this complex on the levels of incorporation of radioactively in macromolecules. Similar assays carried out by us confirmed that the complex Rh(I)(CO)2 Cl (2-Aminobenzothiazole) does not significantly inhibit the synthesis of macromolecules of the parasite, whereas Rh(I)(CO)2 Cl (5-Cl-2-Methylbenzothiazole) inhibited the incorporation of thymidine by approximately 72% after 135 min at 100 mg/ml, and leucine and uridine incorporation by 60% for the same time and dosage. The cytotoxicity of the complex Rh(I), with ligand (2- Aminobenzothiazole) to J-774 macrophages at 24 hr and maximum dosage, produced 24.2% specific liberation of Cr^51,whereas the complex Rh(CO)2 Cl (5-Cl-2-Methylbenzothiazole), which produced the greatest inhibition rates in macromolecule biosynthesis in Leishmania, was 3.2% at 24 hr of maximum dosage. The ultrastructural alterations observed in the promastigote forms of L. donovani after treatment with Rh(CO)2 Cl (5-Cl-2-Methylbenzothiazole) show large abnormal chromatin condensations in the nucleus and kinetoplast of the parasite, which is indicative of an intercalation of the metal complex in the DNA. In addition, numerous vacuoles appear, some with multiple membranes and the intravacuolar appearance of fingerprint myeline figures. In some cases a pronounced swelling of the mitochrondria is visible, with conspicuous mitochondrial crests and electrondense deposits in the matrix. Leishmania promastigotes lose their typical structure and amoebal cytoplasmic projections appear as a consequence of the loss of tone in the tubules which make up the cytoskeleton. In some cases, discontinuities appear in the nuclear membrane and in the lacunar spaces between the nuclear membrane and the cytosol, as well as between the tubules of the cytoskeleton and the cytoplasm. These effects on the membranes could be due to a direct influence of the ligand on the biosynthesis of the membrane components, which would lead to irregularities and dysfunctions. In the present work, we study the manner of action and the cytopathological effects of certain metal complexes which in vitro have shown leishmanicidal activity, seeking alternatives to the traditional medicines for leishmaniasis. The best candidate, given its slight toxicity for mammal cells and its high activity against Leishmania by inhibiting the synthesis of macromolecules, is the complex Rh(I): Rh(CO)2 Cl (5-Cl-2-Methylbenzothiazole). References Bacchi CJ, Nathan HC, Hutner SH, McCan PP, Sjoerlma A 1980. Polyamine metabolism: a potential therapeutic target in trypanosomes. Science 210: 332-334. Bacchi CJ, Sufrin JR, Nathan HC, Spiess AJ, Hannant T, Garofalo J, Alecia K, Katz L, Yarlett N 1991. 5-Alkyl- Substituted Analogs of 5-Methylthio adenosine as trypanocides. Antimicrob Agents Chemother 35: 1315-1320. Balder AE, Gonias SL, Pizzo SV 1985. Trypanosoma brucei gambiense: growth in vitro of bloodstream forms inhibited by dichlorodiamineplatinum(II) compounds and hypolipidemic drugs. Exp Parasitol 59: 74-80. Berman JD 1988. Chemotherapy for leishmaniasis: biochemical mechanisms, clinical efficacy, and future strategies. Rev Infect Dis 10: 560-585. Bujdakova H, Kuchta T, Sidoova E, Gvozdjalova A 1993. Anti- Candida Activity of Four Antifungal Benzothiazoles. FEMS Microbiol Letters 112: 329-334. Castilla-Calvente JJ, Sanchez-Moreno M, Mesa-Valle C, Osuna A 1995. Leishmania donovani: In vitro culture and [^1H]NMR characterization of amastigote-like forms. Mol Cell Biochem 142: 89-97. Craciunescu DG, Furlani A, Scarcia V, Doadrio A 1985. Synthesis, Cytostatic and Antitumor Propierties of New Rh(I) Thiazole Complexes. Biol Trace Elem Res 8: 251- 261. Craciunescu DG, Molina C, Alonso MP, Parrondo-Iglesias E, Doadrio-Lopez A, Doadrio-Villarejo JC, Gaston de Iriarte E, Ghirvu CI, Eecoli N 1990a. Estudio de las actividades farmacologicas duales (antitumorales y antitripanosomicas) y del perfil toxicologico de algunos nuevos complejos del Rh(I) y del Ir(I). An Real Acad Farm 56: 181-200. Craciunescu DG, Molina C, Carvajal RM, Doadrio-Lopez A, Berger M, Ghirvu C 1986. Estudio de las actividades antitumorales y/o antitripanosomicas de dos nuevas series de complejos del Pt(II): cis-Pt(L)^2 Br^2 (Donde L=Derivado de tiazol y/o derivado de sulfonamida) y cis-Pt(L)(X) (Donde n=1 o 2;L=1,2- Diaminociclohexano y X- o X2- = Aniones mono o bidentados de acidos organicos). An Real Acad Farm 52: 45- 64. Craciunescu DG, Molina C, Doadrio A, Parrondo E, Gomez A, Sanchez de Leon M, Gaston de Iriarte E, Ghirvu C 1987. Sintesis, caracterizacion estructural, estudio de las propiedades duales (actividades antitumorales y actividades antitripanosomicas), y evaluacion de la nefrotoxicidad de algunos nuevos complejos RhIII(L)^3, donde L= derivado de alquil o aril ditiocarbamato y/o xantato. An Real Acad Farm 53: 205-232. Craciunescu DG, Molina C, Parrondo-Iglesias E, Alonso MP, Doadrio-Villarejo JC, Gutierrez-Rios MT 1990b. Complejos cationicos del Rh(III) y del Ir(III) con los medicamentos antimalaricos; actividades farmacologicas duales in vivo (antitumorales y antitripanosomicas). An Real Acad Farm 56: 453-468. Craciunescu DG, Parrondo E, Alonso MP, Molina C, Doadrio A, Gomez A, Mosqueira RM, Ghirvu C, Gaston de Iriarte E 1988a. Simntesis, caracterizacion estructural, estudio de las propiedades estructurales (actividades antitumorales y antitripanosomicas) y evolucion de la nefrotoxicidad de algunos nuevos complejos (OsIII(L)^3)^0, y (RuIII(L)^3)^0 donde L=derivado de alquil o aril ditiocarbamato y/o xantato. An Real Acad Farm 54: 16-45. Craciunescu DG, Parrondo E, Gaston de Iriarte E, Ghirvu C 1988b. Estudio de las actividades farmacologicas duales (actividades antitripanosomicas y actividades antitumorales) y de la nefrotoxicidad de algunos nuevos complejos (RuII(L)^2)^0 y (IrII(L)^2)^0 donde L=derivado de alquil o aril ditiocarbamato y/o xantato. An Real Acad Farm 54: 46-69. Croft SL 1988. Recent developments in the chemotherapy of leishmaniasis. Trends Pharmacol Sci 9: 376-381. Croft SL, Neal RA, Craciunescu DG, Certad-Fombona G 1992. The activity of Pt, Ir and Rh drug complexes against L. donovani. Trop Med and Parasitol 43: 24- 28. Dehand J, Jordanov J 1976. Complexes of platinum (II) with substituted thiazolines. Chem Biol Interact 11: 605-610. Englund PT, Hajduk L, Marini JC 1982. The molecular biology of trypanosomes. Annu Rev Biochem 51: 695-698. Farrell NP, Williamson J, McLaren JM 1984. Trypanocidal and antitumor activity of platinum metal and platinum-metal-drug dual function complexes. Biochem Pharmacol 33: 961-971. Fouce RB, Escribano JM, Alunda JM 1991. Putrescine Uptake Regulation in Response to Alpha-Difluoromethylornithine Treatment in Leishmania-infantum Promastigotes. Mol Cellular Biochem 107: 127-133. Fulfor DE, Bradley SG, Marcino-Cabral F 1986. Cytopathogenicity of Naegleria fowleri for cultured Rat Neuroblastoma Cells. J Protozool 32: 176- 179. Goad LJ, Berens RL, Marr J, Beach DH, Holz Jr GG 1989. The activity of ketoconazole and other azoles against Trypanosoma cruzi. Biochemistry and chemotherapeutic action in vitro. Mol Biochem Parasitol 32: 179-190. Goad LJ, Holtz GG, Beach DH 1985. Sterols of ketoconazole- inhibited Leishmania mexicana mexicana promastigotes. Mol Biochem Parasitol 15: 257-279.
Gonzalez AM, Castanys S, Osuna A, Herrador MM, Dominguez J 1989. Inhibitory Effect of New Pyrimidine Bases on Trypanosome cruzi. Arch Pharm (Weinheim) 332: 843-846. Haugwitz RD, Angel RG, Jacobs GA, Maurer BV, Narayanan VL, Cruthers LR, Szanto J 1982. Antiparasitic agents. Synthesis and antihelmintic acitivities of novel 2-heteroaromatic- substituted isothiocyanatobenzoxazoles and benzothiazoles. J Med Chem 25: 969-974. Hisano T, Ichikawa M, Tsumoto K, Tasaki M 1982. Synthesis of benzoxazoles, Benzothiazoles and benzimidazoles and evaluation of their antifungal, insecticidal and hervicidal activities. Chem Pharmacol Bull 30: 2996-3004. Kinnamon KE, Steck EA, Rane DS 1979. Activity of antitumor drugs against African trypanosomes. Antimicrob Agents Chemother 15: 157-160. Marr JJ 1991. Purine analogs as chemotherapeutic agents in Leishmaniasis and American Trypa-nosomiasis. J Lab Clinical Med 118: 111-119. Mesa-Valle CM, Craciunescu D, Parrondo-Iglesias E, Osuna A 1989. In vitro Action of Platinum (II) and Platinum (IV) Complexes in Trypanosoma cruzi and Leishmania donovani. Arzneimittel Forschung 39: 838- 842. Mesa-Valle CM, Moraleda-Lindez V, Craciunescu D, Alonso MP, Osuna A 1993. In vitro Action of New Organometallic Compounds agaisnst Tripanosomatidae protozoa. Arzneimittel Forschung 43: 1010-1013. Modabber F 1987. A Global Partnership, 8th Programme report, p. 99-112. In J Maurice, AM Pearce, Tropical Disease Research, WHO, Geneva. Opperdoes FR, Baudhuin P, Coppens J, De Roe C, Edwards SW, Weijers PJ, Misset O 1984. Purification, morphometric analysis, and charecteri-zation of the glycosomes (microbodies) of the protozoan hemoflagellate Trypanosoma brucei. J Cell Biol 98: 1178-1184. Osuna A, Castanys S, Ortega G, Gamarro F, Aneiros J, Brana MF, Roldan CM 1983. Estudio ultraestructural de la accion de dos derivados benzo(de) isoquinolil-1,3-diona sobre T. cruzi. Rev Inst Med Trop Sao Paulo 25: 133- 138. Ruiz-Perez LM, Osuna A, Castanys S, Gamarro F, Craciunescu D, Doadrio A 1986. Evaluation of the toxicity of Rh(III) and Pt(II) complexes against T. cruzi culture forms. Arzneimittel Forschung 36: 13-16. Ruiz-Perez LM, Osuna A, Lopez MC, Castanys S, Gamarro F, Craciunescu D, Alonso C 1987. Mode of action of intercalating drug, cis-Pt(II)(DDH) Cl2,cis-Pt(II)(DDH) (metafluoro- benzoic)^2 and cis-Pt(II)(DDH) (mucobromic)^2 on T. cruzi. Trop Med Parasitol 38: 45-48. Sanchez-Delgado R, Lazardi K, Rincon L, Urubina JA 1993. Toward a Novel Metal-Based Chemotherapy against Tropical Diseases. 1. Enhancement of the Efficacy of Clotrimazole against Trypanosome cruzi by Complexation to Ruthenium in RuCl^2(Clotrimazole)^2. J Med Chem 36: 2041- 2043. Sjoerdsma A, Schechter PJ 1984. Commentary: Chemotherapeutic implications of polyamine biosynthesis inhibition. Clin Pharmacol Ther 35: 287-300. Urbina JA, Maldonado RA, Payares G, Marchan E, Lazardi K, Aguirre T, Piras MM, Piras R, De Souza W 1991. In vitro and in vivo synergism of binary and ternary combinations of ergosterol biosynthesis inhibitors as antiproliferative agents against Trypanosoma cruzi: Therapeutic implications. Mem Inst Oswaldo Cruz 86: 33-35. WHO 1984 Technical Report Series. The Leishmaniases. 701 Willson M, Perie JJ, Malecaze F, Opperdoes F, Callens M 1992. Biological properties of amidinium sulfinic and sulfonic acid derivatives: inhibition of glycolytic enzymes of Trypanosoma brucei and protective effect on cell growth. Eur J Med Chem 27: 799-808. Wysor MS, Swelling LA, Sanders JA, Grenan MM 1982. Cure of mice infected with Trypanosoma rhodesiense by cis- diamine dichloroplatinum II and disulfiram rescue. Science 217: 454-456. Yalcin I, Sener E 1993. QSARs of some novel antibacterial benzimidazoles, benzoxazoles, and oxazolo-pyridines against an enteric gram-negative rod K. pneumoniae. Intern J Pharmaceutics 98: 1-8. Zahner H, Franz M, Kohler P, Striebel HP 1991a. Antifilarial Efficacy In vitro of 2-tert-Butyl-Benzothiazole Derivatives on Adult Litomosoides Carinii. ArzneimittelForschung 41: 821-827. Zahner H, Johri GM, Kohler P, Striebel, Franz M 1991b. In vitro Effects of 2-tert-Butyl-Benzothizaole Derivatives on Microfilariae of Litomosoides Carinii, Brugia Malayi and Acanthocheilonema Viteae. ArzneimittelForschung 41: 764-768. Zinsstag J, Brun R, Craciunescu DG, Parrondo-Iglesias E 1991. In vitro activity of organometallic complexes of Ir, Pt and Rh on Trypanosoma b. gambiense, T.b. rhodesiense and T.b. brucei. Trop Med Parasitol 42: 41-44. --------------------------------------------------------------
Fig. 2: effect of Rh^II(CO)2Cl (2- Aminobenzothiazole) on the incorporation by Leishmania donovani of: (A).- (^3H)leucine, (B).- (^3H)thymidine, and (C).- (^3H)uridine. (l) Control; (n) Drug concentration-100 mg/ml; (s) Drug concentration-50 mg/ml. Fig. 3: effect of Rh^II(CO)2Cl (5-Cl-2- Methylbenzothiazole) on the incorporation by Leishmania donovani of: (A).- (^3H)leucine, (B).- (^3H)thymidine, and (C).- (^3H)uridine. (l) Control; (n) Drug concentration-100 mg/ml; (s) Drug concentration-50 mg/ml. Fig. 4: ultrastructural effects of cis- Pt^II (DDH)(2,5-Dihidroxy-benzensulphonic)^2 of Leishmania donovani in vitro Nucleus (N), Kinetoplast DNA (K), Mitochondria (M). (A) (x7.000) and (B) (x12.000) Fig. 5: Leishmania exposed for 8 hr in vitro to 100 mg/ml Rh^II(CO)2Cl (5-Cl-2- Methylbenzothiazole). Nucleus (N). Mitochondria (M). Kinetoplast DNA (K). (x15.000). Fig. 6: section showing an normal Leishmania (x12.000). Nucleus (N). Mitochondria (M). Kinetoplast DNA (K).
--------------------------------------------------------------
TABLE I Percentage of growth inhibition caused by the
metal complexes in the culture of Promastigote and Amastigote
like forms of Leishmania donovani
--------------------------------------------------------------
Time of incubation
24 hr 48 hr 72 hr
Concentration --------------- -------------- --------------
(mg/ml) 100 10 1 100 10 1 100 10 1
--------------------------------------------------------------
Promastigote forms
Rh(CO)2Cl 54.9 0 0 60 0 0 97 0 0
(2-Aminobenzothiazole)
Rh(CO)2Cl 72.5 0 0 64.4 26.4 0 83.6 14.2 0
(5-Cl-2-Methilbenzothiazole)
Cis-Pt(DDH) 43.3 0 0 47.8 26.3 0 67.2 37 0
(2,5-Dihidroxibenzensulphonic)^2
Amastigote like forms
Rh(CO)2Cl 67.8 0 0 72.7 0 0 86.5 0 0
(2-Aminobenzothiazole)
Rh(CO)2Cl 31.9 0 0 31 0 0 50.8 0 0
(5-Cl-2-Methilbenzothiazole)
Cis-Pt(DDH) 52.7 24.7 0 50.1 29.7 0 57.8 18.6 0
(2,5-Dihidroxibenzensulphonic)^2
The results are the average of five experiments.
--------------------------------------------------------------
TABLE II Cytotoxic effect of metal complexes on J-774
Macrophages^a
--------------------------------------------------------------
Time of incubation
24 hr 48 hr 72 hr
Concentration ------------------ -------------- ---------
(mg/ml) 100 10 1 100 10 1 100 10
--------------------------------------------------------------
Rh(CO)2Cl 0 0 0 2.4 3.2 0 24 3.2
(2-Aminobenzothiazole)
Rh(CO)2Cl 0 0 2.9 0 0 3.2 0
(5-Cl-2-Methilbenzothiazole)
Cis-Pt(DDH) 0 0 0 3.5 0 0 3.0 0
(2,5-Dihidroxibenzensulphonic)^2
a: the results are the averages of five different experiments,
expressed in percentage of specific liberation of Cr^51.
--------------------------------------------------------------
Copyright 1996 Fundacao Oswaldo Cruz
The following images related to this document are available:Photo images[oc96113f.jpg] [oc96113e.jpg] [oc96113d.jpg]Line drawing images[oc96113c.gif] [oc96113a.gif] [oc96113b.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}