 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memorias do Instituto Oswaldo Cruz RESEARCH NOTE Slit Cuticular Structure Observed by Scanning Electron Microscopy: Presence on the Walking Legs of Amblyomma cajennense(Fabricius,1787) (Acari: Ixodidae) Katia Maria Famadas/+/++, Reinalda Marisa Lanfredi */++,Nicolau Maues da Serra-Freire **/++
Laboratorio de Ixodides, Departamento de Entomologia,
Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ,
Brasil
+Corresponding author. Fax: +55-21-290.9339
Received 24 November 1995
Code Number: OC97011
Sizeof Files:
Text: 8.9K
Graphics: Line drawings (gif) - 175.1K
Tables (gif) - 34.4K
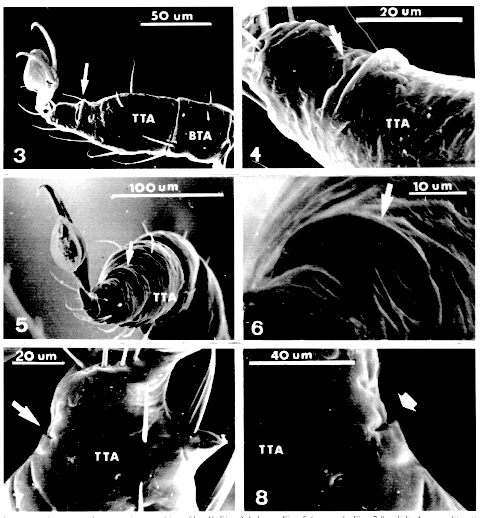
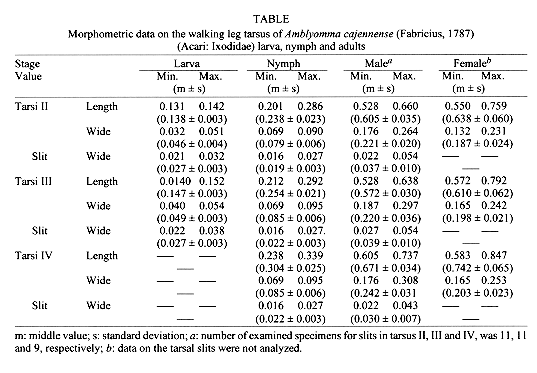
Key words: Amblyomma cajennense - tick - slit cuticular structure - distal tarsal slit sense organ - lyrifissure - scanning electron microscopy - Ixodidae RESEARCH NOTE Several types of cuticular structures on ticks have been observed by scanning electron microscopy (SEM) (TA Wooley 1972 TransAmer MicrosSoc 91: 35-47, E Hess & M Vlimant 1984 p. 253-260. InDA Griffiths & CE Bowman (eds) Acarology, Ellis Horwood Ltd,Chichester, UK, KM Famadas et al. 1984 Mem Inst Oswaldo Cruz 89:571-573). One of them, described by Hess and Vlimant (loc. cit.),was named the distal tarsal slit sense organ (DTSSO) since exteriorly it resembles the slit sense organ of spiders. The common house spider has atarso-metatarsal lyriform organ on each of its eight legs (CWalcott 1969 Am Zoologist 9: 133-144). Slits were also described on the legs of adult ticks of Ixodesricinus (Linnaeus), mainly associated with eudesmatic or adesmatic articulations (L Van der Hammen 1983 ZoologischeMeded Leiden57: 209-242). Since the first description of the slit in spiders by Bertkau (1878, in JWS Pringle 1955 J Exp Biol 32: 270-278) different terms have been used to designate it. The term lyriform organ has been used to describe a compound organ of spiders, it is marked mainly by a group of slits. Single slits are common in Acari, referred to as lyrifissures (Vander Hammen loc. cit., TA Wooley 1988 Acarology - Mites and human welfare J Wiley & Sons, New York, USA 484 pp.). Although slits occur in nearly all Chelicerata (NE M'Indoo1911 Proc Acad Natur Sci Philadelphia 63: 375-418, BJ Kaston 1935 J Morphol 58: 189-209, Pringle loc. cit. ), most studies on this structure are made from Aranea. Recent advances in electrophysiology, electron microscopy and experimental models with spiders have shown many sensorial functions attributed to this organ, also indicating that the slit acts as a mechanoreceptor sensitive to pressure upon the cuticle (EA Seyfarth & FG Barth 1972J Comp Physiol 78: 176-191, fg Barth 1972 J Comp Physiol 78: 315-336), or vibration receptor (Walcott loc. cit.).Seyfarth and Barth (loc. cit. ) and Barth (loc. cit. ) have proved its role on the kinetic direction. Besides these functions, Hessand Vlimant (loc. cit.) supposed that it could also play a role asa gravity receptor in ticks. Knowledge of a new sense structure of ticks is an important step for the development of better methods, based on the ticks' need to find suitable feeding host. The aim of this paper is to record the presence of slits on the dorsal surface of telotarsus II, III, IV in larvae, nymphs and adults of Amblyomma cajennense (Fabricius, 1787). Engorged larvae and nymphs of A. cajennense, collected on horses, were kept under laboratory conditions (25-29 C, 80+/-10% RH), until ecdysis. Larvae aged 15 days, nymphs and adults for 24 hr after hatching, unfed, were killed and preserved in 70% ethanol Famadas et al. loc. cit.). A sample of 30 larvae, 30 nymphs, 20 males and 20 females ofA. cajennense was prepared for optical microscopy (MO) according to KM Famadas' (1993 Universidade Federal Rural do Rio de Janeiro, RJ, Brasil xx + 86pp.; 1996 Mem Inst Oswaldo Cruz 91: 139-140) method. Another batch of 20 larvae, 10 nymphs, 5 males and 5 females was prepared for SEM using the method of JE Keiranset al. (1976 Acarologia 18: 217-225). The electron micrographies were carried out on a Stereoscan 200 SEM at 16Kv. All measurements are expressed in millimeters. The average is followed by standard deviation and the variation interval, representing a sample of right-legs II-IV of specimens measured on a OlympusBH2 opticalmicroscope. In order to compare regression and correlation means, T tests were applied. A slit on the dorsal telotarsus face of walking legs of larvae, nymphs and adults of A. cajennense was observed through MO and SEM. Although Hess and Vlimant (loc. cit.) only have depicted the adult slit of A. variegatum (by SEM micrography), based on their description of DTSSO, we think it would be the same structure in A. cajennense.
The slit can be easily observed through optical microscopy (200x)
in larvae (
As also observed by M'Indoo (loc. cit.) and Kaston( loc.cit.),
in Arachnida, the slits in each stage of A.cajennense are located on
a slight elevation of the cuticle Figs 1-2and Figs.3-8). In MO it is seen as
a thick orange brown area darker than other areas (Figs 1, 2). This slit is
constituted of a larger proximal and distal border (Fig. 2) and it is
arranged crosswise to the segment axis. Through SEM, in dorsal and lateral
views,the slit resembles a mouth (Figs 3-6).
Figures 3-8
Amblyomma cajennense, electron micrographies of leg II.
The size of the slit seems relatively constant about the taxa up to
now reported (Ixodes ricinus, Van der Hammen
loc.cit.;A. variegatum, Hess & Vlimant loc.cit.;
spiders,Kaston loc. cit.).
The slits present on the tibia and femur of spiders legs have an
important role in the kinestethic orientation and they are
specifically adapted to change the stimulus on cuticle surface deformations
into a stimulus carried via sense cells. In addition the deformation of
a single slit is greatest in its middle region and decreases towards its
ends; the deformability of the slit increases according to its
length (Barth loc. cit.).
Acknowlegments: to Dr Sergio M Faria, National Research Center of
Agrobiology (CNPA), Empresa Brasileira de Pesquisa Agropecuaria (EMBRAPA),
Seropedica, Rio de Janeiro, Brazil, for utilization of the scanning electron
microscope.
Copyright 1996 Fundacao Oswaldo Cruz
|
| |||||||||

{kind=link}
{kind=link}
{kind=link}