 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
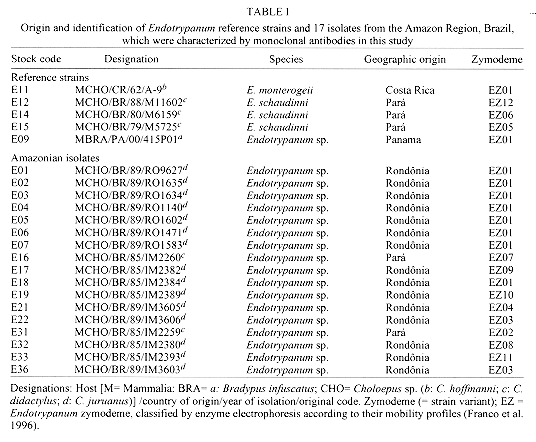
Vol. 92(1), Jan./Feb. 1997 Characterization of Endotrypanum Parasites Using Specific Monoclonal Antibodies Antonia Maria Ramos Franco/+, Gerzia MC Machado, Roberto D Naiff*, Celia FS Moreira, Diane McMahon-Pratt**, Gabriel Grimaldi Jr
Departamento de Imunologia, Instituto Oswaldo Cruz, Av. Brasil 4365,
21045-900 Rio de Janeiro, RJ, Brasil +CAPES fellowship and corresponding author. Fax: +55-21-280.1589 Received 9 April 1996. Accepted 21 October 1996
Code Number:OC97013
Sizes of Files:
Text: 22.2K
Graphics: Photographs (jpg) - 37.4K
Tables (gif) - 156.1K
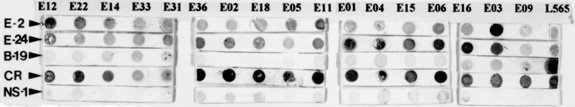
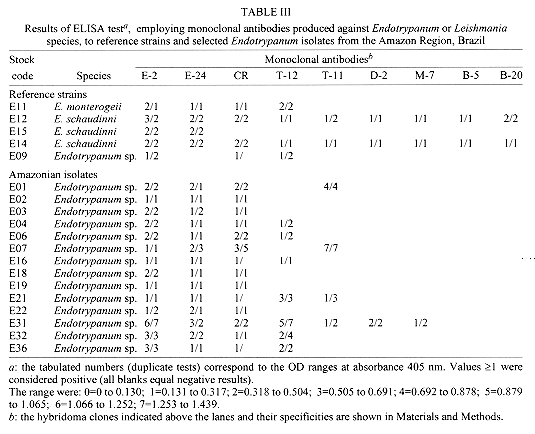
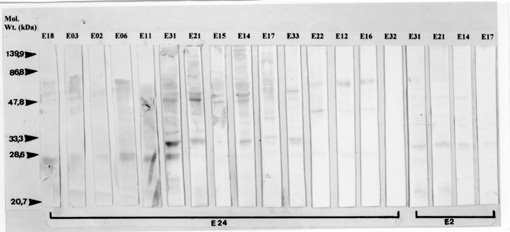
A large number of Endotrypanum stocks (representing an heterogeneous population of strains) have been screened against a panel of monoclonal antibodies (MAbs) derived for selected species of Endotrypanum or Leishmania, to see whether this approach could be used to group/differentiate further among these parasites. Using different immunological assay systems, MAbs considered specific for the genus Endotrypanum (E-24, CXXX-3G5-F12) or strain M6159 of E. schaudinni (E-2, CXIV-3C7-F5) reacted variably according to the test used but in the ELISA or immunofluorescence assay both reacted with all the strains tested. Analyses using these MAbs showed antigenic diversity occurring among the Endotrypanum strains, but no qualitative or quantitative reactivity pattern could be consistently related to parasite origin (i.e., host species involved) or geographic area of isolation. Western blot analyses of the parasites showed that these MAbs recognized multiple components. Differences existed either in the epitope density or molecular forms associated with the antigenic determinants and therefore allowed the assignment of the strains to specific antigenic groups. Using immunofluorescence or ELISA assay, clone E-24 produced reaction with L. equatorensis (which is a parasite of sloth and rodent), but not with other trypanosomatids examined. Interestingly, the latter parasite and the Endotrypanum strains cross-reacted with a number of MAbs that were produced against members of the L. major-L. tropica complex. Key words: Endotrypanum - Kinetoplastida - Trypanosomatidae - mammalian reservoirs - molecular characterization - monoclonal antibodies Endotrypanum parasites are unique among the Kinetoplastida in that they infect erythrocytes of their mammalian host, the two and three-toed sloths (genera Choloepus and Bradypus, respectively). Inside the erythrocyte the Endotrypanum assumes an epimastigote or trypomastigote form, while in the sandfly and during in vitro culture the parasite assumes promastigote morphology (Shaw 1992). Similarities at the morphological, molecular and biological levels exist between many trypanoso-matids isolated from sylvatic insect and/or vertebrate reservoir hosts that make the identification of the medical importance parasites demanding (Shaw 1985). Moreover, the neotropical tree sloths, which are reservoir of at least six Leishmania species pathogenic for human (review in Grimaldi & Tesh 1993), are hosts of Endotrypanum (reviewed in Shaw 1992) and several Trypanosoma species [T. cruzi, T. rangeli, T. leeuwenhoeki, T. preguici and T. legeri] (Montero-Gei 1956, Shaw 1969, Zeledon et al. 1975, Herrer & Christensen 1980, Miles et al. 1983). Identification of Endotrypanum strains usually relies on the examination of promastigotes from cultures, and those forms of the parasite are morphological indistinguishable from Leishmania (Croft et al. 1980). More precise taxonomic markers for Endotrypanum, however, have resulted from the application of molecular techniques (Greig et al. 1989, Lopes et al. 1990, Fernandes et al. 1993, Medina-Acosta et al. 1994, Franco et al. 1996). Among the new approaches for identifying Endotrypanum is serodeme analysis using specific MAbs (Lopes & McMahon-Pratt 1989). In order to extend these observations, here we discuss the results of parasite differentiation of Endotrypanum (comprised of an heterogenous population of reference strains and Amazonian isolates), as characterized by their reactivities with MAbs and using distinct assay systems. MATERIALS AND METHODS The parasites - The origin (i.e., host species involved or geographical area of isolation) of the 22 Endotrypanum stocks examined in this study, which included reference strains (Medina-Acosta et al. 1994) are given in Table I. The stocks were stored as stabilates at -190oC in liquid nitrogen refrigerators, and the freezing medium used was Schneider's medium containing 10% fetal bovine serum (FBS) and 8% dimethylsulfoxide (Grimaldi et al. 1992). Preparation of samples - Procedures for growing Endotrypanum promastigotes in vitro and for preparation of extracts for serodeme analysis with monoclonal antibodies have been reported previously (McMahon-Pratt et al. 1986). Briefly, promastigotes in the log phase of growth in Schneider's medium were harvested by centrifugation (1,500 x g for 10 min at 4 C) and washed twice in phosphate-buffered saline (PBS), pH 7.3. The final pellet was used for preparation of samples. Antigens of each of the strains tested were prepared as promastigote homogenates containing protease inhibitors (Leon et al. 1992). Prior to analysis, the samples were briefly sonicated using a bath sonicator (RAI Research, Model 250 Ultrasonic cleaner) to homogenize the antigens, and then centrifuged (2,000 x g for 10 min, at 4 C). Protein concentration was measured by the protein assay method of Bradford (1976), and all samples were resuspended to the same concentration before analysis. Monoclonal antibodies and immunological assay systems - The MAbs used in this study were produced against membrane components of Endotrypanum (Lopes & McMahon-Pratt 1989) or Leishmania species (McMahon-Pratt et al. 1986). The following group- or species-specific clones were employed for typing parasites: E-24, CXXX-3G5-F12, Endotrypanum sp.; E-2, CXIV-3C7-F5, E. schaudinni; T-11, LXIX 2D8-D7, L. (L.) major; T-12, IS1 2G7-F1, L. (L.) tropica; D-2, LXXVIII 2E5-A8, L. donovani complex; M-7, IXVIII 1D7-B8, L. mexicana complex; B-20, 2 2F7-D3, Leishmania (V.) sp.; B-5, VII-2C5-C5, L. braziliensis complex; B-19, XLIV-5A2-B9, L. (V.) guyanensis; CR, G2G4, cross-reactive to all kinetoplastids (Lopes & McMahon-Pratt 1989, WHO 1993, Grimaldi & McMahon-Pratt 1996). Typing of the Endotrypanum with MAbs was performed with an indirect immunofluorescence assay (IFAT), using whole fixed promastigotes as antigen (McMahon-Pratt et al. 1986) or by distinct immune binding assay systems such as indirect radioimmune assay (RIA) (Grimaldi et al. 1992), the ELISA technique (Jaffe & McMahon-Pratt 1987), or dot-blot ELISA (Hawkes et al. 1982), using whole parasite lysates as antigen (Grimaldi et al. 1992). Characterization of the molecules associated with the specific antigenic determinants was performed by Western blot analysis (Jaffe & McMahon-Pratt 1983). Briefly, the soluble antigen extracts were resolved by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) using 10% slab gels in non-reducing conditions, and electrophoretically transferred to nitrocellulose paper (Schleicher and Schuell, Keene, New Hampshire, USA). The techniques have been described in detail previously (Leon et al. 1992). RESULTS AND DISCUSSION In this paper, the characterization of representative strains of Endotrypanum, based on their reactivity patterns using specific MAbs to Endotrypanum (Lopes & McMahon-Pratt 1989) or Leishmania (WHO 1993, Grimaldi & McMahon-Pratt 1996) is presented. The genus Endotrypanum, analyzed in this study (Table I), may represent an heterogeneous complex of parasite species or strain variants, as classified by numerical zymotaxonomy (Franco et al. 1996). Reactivity with monoclonal antibodies -
The analysis showed antigenic variation occurring among the Endotrypanum strains. Significant differences between the reactive patterns with specific MAbs could be observed among stocks ( Table II, Table III), but no qualitative or quantitative pattern could be consistently related to parasite origin (i.e., host species involved) or geographical area of isolation. These differences can be related with strain variation in the level of expression of certain antigenic determinants, as recognized by these MAbs. Variation in the sensitivity of the test may also occur due to the type of screening assay used. IFAT studies were carried out to confirm the specificity and binding of the MAbs to the Endotrypanum surface determinants on intact parasites (Table IV). Each clone reacted in a continuous pattern on the surface of the promastigotes. Interestingly, clone E-24 produced the same immunofluorescent pattern of reaction (data not shown) with a parasite of sloth and rodent, L. equatorensis (Grimaldi et al. 1992). As noted with L. colombiensis (Kreutzer et al. 1991), two strains of L. equatorensis cross-reacted with MAbs T1 (clone XLVI-5B8-B3), T2 (XLVI, 4H12-C2), T3 (XLVI-5A5-D4), T4 (LXVIII-1A4-G1) and T8 (LXVII-3E12-F8) (Grimaldi et al. 1992), which were originally produced against parasites in the L. major/L. tropica complex (Jaffe & McMahon-Pratt 1983). Moreover, using either IFAT (data not shown) or ELISA assay (Table III) the Endotrypanum strains reacted with MAbs specific to Leishmania, particularly with clones IS1-2C8-C7 (T-12) and LXIX-2D8-D7 (T-11) which are L. major- or L. tropica-specific respectively (WHO 1993). Our data confirm a previously reported study (Shaw 1992) showing that a number of Endotrypanum strains cross-reacted with these MAbs (Jaffe & McMahon-Pratt 1983) or monoclonal antibody (WIC 79.3) which recognizes lipophosphoglycan components of L. major (Handman et al. 1984). Based on this results, it appears that close antigenic links may exist between Endotrypanum and Leishmania. Work is now in progress to better define the phylogenetic relationship between these parasites. Whatever the explanation for the existence of these cross-reactive epitopes, these results indicate caution should be taken by all researchers working with field isolates. Identification of Endotrypanum antigens by MAbs -
In conclusion, we have classified a total of 17 Amazon Endotrypanum isolates (from the states of Rondonia and Para) through the application of techniques employing specific MAbs in comparison with standard reference strains. Characterization of Endotrypanum with specific MAbs is an additional approach useful to group/differentiate further among these parasites, but problems related to the identification of stocks (the described species are indistinguishable) were encountered using the MAbs employed in this study. Our results confirm previous studies (Lopes et al. 1990, Franco et al. 1996) reporting heterogeneity in this genus. ACKNOWLEDGMENTS To Dr JJ Shaw (Evandro Chagas Institute, PA, BR) for the donation of reference strains. REFERENCES
This work was supported in part by the Brazilian National Council of Scientific Development and Technology (CNPq) and CAPES/BR.
Copyright 1996 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97013f.jpg] [oc97013c.jpg]Line drawing images[oc97013e.gif] [oc97013d.gif] [oc97013b.gif] [oc97013a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}