 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 92(1), Jan./Feb. 1997 RESEARCH NOTE Invasion and Development of Trypanosoma cruzi in Primary Cultures of Mouse Embryo Hepatocytes R Porrozzi, R Soares, M Meuser, C Guguen-Guillouzo*, MNL Meirelles/+
Laboratorio de Ultra-estrutura Celular, Departamento de
Ultra-estrutura e Biologia Celular, Instituto Oswaldo Cruz, Av. Brasil
4365, 21045-900 Rio de Janeiro, RJ, Brasil
+Corresponding author. Fax: +55-21-260.4434
Received 19 April 1996
Code Number:OC97025
Sizes of Files:
Text: 11.6k
Graphics: Line drawings (gif) - 3.7K
Photographs (jpg) - 101.4K
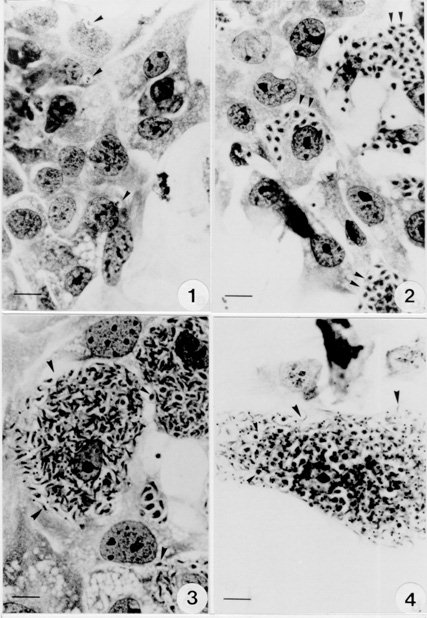
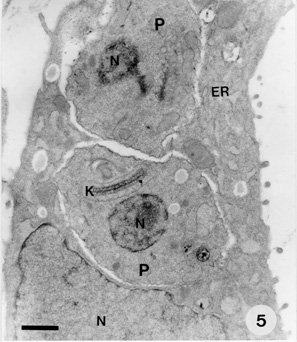
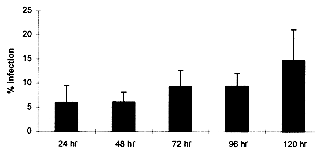
Key words : Trypanosoma cruzi - cell culture -hepatocytes - polarized cells RESEARCH NOTE Trypanosoma cruzi, the aetiological agent of Chagas' disease, has to invade a vertebrate host cell for the necessary fulfilment of its complex life cycle. Despite the extensive efforts to characterize the molecular bases of the parasite pathogenicity, many questions remain unanswered, although considerable progress has been made to understand the mechanism of parasite invasion and survival within the host cells (JA Dvorak & TP Hyde 1973 Exp Parasitol 34: 268-283, N Nogueira & ZA Cohn 1976 J Exp Med 143: 1402-1420, B Zingales et al. 1982 Mol Biochem Parasitol 6: 111-124, MNL Meirelles et al. 1984 J Submicrosc Cyt 16: 533-545, MNL Meirelles et al. 1986 Eur J Cell Biol 41: 198-206, W De Souza 1989 Progress in Protistol 3: 87-184, S Schenkman et al. 1988 Cell 55: 157-165, I Tardieux et al. 1994 Cell 71: 1117-1130, SNJ Moreno et al. 1994 J Exp Med 180: 1535-1540, R Do Campo & SNJ Moreno 1996 Parasitol Today 12: 61-65). T. cruzi is predominantly found infecting myocardial cells, macrophages, and cells of the autonomic nervous system of its mammalian host (Z Brener 1973 Ann Rev Microbiol 27: 347-382). The parasite can invade many cell types in vitro, including epithelial cells in a polarized manner (Schenkman et al . loc. cit.). The tropism of T. cruzi for different tissues, including liver, has also been reported (RC Melo & Z Brener 1978 J Parasitol 64: 475-482, N Deutshlander et al. 1978 Tropenmed Parasitol 29: 323-326). However, the target cells in these studies seem to be tissue resident macrophages. Hepatocytes infections with T. cruzi have been reported in association with bacteria in immunosuppressed mice (KS Calabrese et al. 1991 Immunol Lett 31: 91-96). We are now reporting T. cruzi invasion and development within an in vitro system of primary culture of mouse embryo hepatocyte. This point to a reproducible system to study the interaction between T. cruzi and polarized cells. Hepatocytes were isolated according to MA Sells et al. (1985) In vitro 21: 216-220, with some adaptations. Briefly, ten mouse embryo livers (weight 1-2 g) were asseptically removed. The livers were washed with Hank's balanced salt solution Ca++ and Mg++ free (HBSS-CMF). Longitudinal cuts were made in each lobe for enzyme access and the livers were then incubated at 37 C for 20 min with 0.05% collagenase (Type II Wortington) in approximately 50 ml HBSS-CMF. The organs were dispersed by pipetting and liver cells were collected by centrifugation (1000 rpm/3 min). Viable cells were purified by sedimentation at room temperature for 10 min with MEM/199 medium containing 10% Fetal Calf Serum (FCS). Cells yield was 2x10^7/ml (total 10 ml) and viability greater than 90%, as assessed by the trypan blue exclusion test. The cells were seeded on plastic Petri dishes and glass coverslips coated with gelatin. Four hours later fresh defined medium (30% M199 : 70% MEM) supplemented with 5mM CaCl2, 10 mg/ml Insulin, Transferrin and Selenium (ITS), 1 ng/ml Glucagon, 50 ng/ml Epidermal growth factor (EGF), 3.5x10^-6M hydrocortisone and 1 mg/ml Bovine Serum Albumin (BSA) was added and replaced daily. The cultures were incubated at 37#176;C in a humidified atmosphere containing 5% CO2. Trypomastigotes of T. cruzi, Y strain, were collected from the blood of infected mice in the peak of parasitemia, purified by differential centrifugation, washed with PBS and adjusted to a 10:1 parasite-cell ratio. The primary cultures infected 24 hr and/or 48 hr after plating, either at 1x10^6 parasites/well in 24 wells plates or 5x10^6 parasites/Petri dishes (35 mm). After 24 hr, the cultures were washed to eliminate free-swimminng parasites and observed for five days. The coverslips were fixed with Bouin's fixative at different time intervals and stained with Giemsa in order to follow the infection by light microscopy. Two independent observers examined at least 200 randomly selected cells at 1000X magnification. The percentage of hepatocytes containing parasites in to the cytoplasm was determined. The interaction assays were carried out in triplicate under identical conditions. Some infected cultures were also fixed for routine transmission electron microscopy. They were washed in PBS and fixed with 2.5% glutaraldehyde in 0.1M cacodilate buffer for 1 hr at 4 C, washed 3 times in the same buffer and fixed with 1% osmium tetroxide (OsO4) in 0.1M cacodilate buffer for 1 hr at 4 C. The cells were washed in buffer, removed with a cell scraper, and then the suspension was dehydrated in graded acetone and embedded in Epon. Ultrathin sections were collected in cooper grids, stained with uranyl acetate and lead citrate and observed with a Zeiss EM-10C transmission electron microscope. Liver cells obtained by collagenase digestion retained membrane molecules for cell-cell interaction. Few minutes after seeding we could observe the formation of cell clusters, prior to attachment. These clusters, containing about 10 cells, strongly attached to the plastic or gelatin substrate after 1 or 2 hr of incubation. From day 1 through day 3 the cells underwent division and several mitotic figures were observed. Hepatocytes cords were recognized and cells displayed an epithelial shape and a polarized organization. However, the nuclei still showed a dispersed chromatin, characteristic of non-differentiated hepatocytes. Thereafter, the cells did not divide and very well differentiated hepatocytes could be observed. The interaction of T. cruzi with hepatocytes was examined in this study using light and electron microscopy. The parasites were often found adhered to the peripheric cells of the clusters. Twenty-four hours after infection, parasites close to the hepatocyte nucleus could be observed by light microscopy (Fig. 1). After 48-72 hr intense parasite proliferation and differentiation was observed (Fig. 2). Differentiation of T. cruzi to try-pomastigote forms was found mostly at 72 and 96 hr and the entire cell-cycle could be accomplished in the hepatocyte primary culture (Figs 3, 4). The ultrastructural observation of the cells 48 hr after infection showed amastigote forms in the hepatocyte cytoplasm (Fig. 5). A quantitative analysis of the infection percentual resulted in values of 6.0+/-3.5%, 6.1+/-2.1%, 9.3+/-3.4%, 9.4+/-2.7% and 14.8+/-6.4%, respectively at 24, 48, 72, 96 and 120 hr of interaction (Fig. 6).
Fig. 1: 24 hr infected hepatocyte displaying two parasites near the cell nucleus. Fig. 2: a great proliferation of the parasites was observed in the cell cytoplasm 48 hr after infection. Figs 3, 4: hepatocyte cells fulled with differentiated trypomastigote forms of T. cruzi. Bar = 20 mm. Figure 5: 48 hr Trypanosoma cruzi -infected hepatocyte showing a well developed endoplasmic reticulum with two amastigote forms in the cytoplasm. ER = endoplasmic reticulum; N = nucleus; P = parasite; K = kinetoplast. Bar = 2 mm. Fig. 6: percentage of infected hepatocytes (day one to day five). Vertical lines indicate standard deviations.
Polarized epithelial cells, like hepatocytes and Madin-Darby canine kidney (MDCK) cells have two distinct plasma membrane domains, the apical and the basolateral, that bear distinct membrane molecules. In a monolayer, basolateral membrane domains are the regions that will make the initial contact between cells and are also associated to increased membrane activity. Studies of T. cruzi invasion of MDCK cells showed that try-pomastigotes enter preferentially through the basolateral surface. Treatment of these cells with citochalasin D selectively inhibited the capacity of the cells to perform endocytosis when membrane-bound and fluid-phase markers were applied to the apical surface, without affecting the endocytosis at the basolateral surface (Schenkman et al. loc. cit.). Our observations with primary culture of hepatocyte cells indicated that within the first hours of interaction, when the cultures were not confluent, the parasites could invade the cells at the basolateral edges. The complete intracellular cycle of the parasite was followed in these cells. It is known that the main target cells for T. cruzi invasion in vivo are macrophages, neuronal and muscle cells. The identification of another cell type that allows invasion by T. cruzi may result in an useful system to identify cell surface components recognized by the parasite during cell invasion.
Supported by CNPq, Papes/FIOCRUZ, PADCT/CNPq, INSERM (U49) Copyright 1996 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97025f6.jpg] [oc97025f5.jpg] [oc97025f1-4.jpg] [oc97025b.jpg] [oc97025a.jpg]Line drawing images[oc97025c.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}