 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Vol. 92(1), Jan./Feb. 1997 Morphology and Histology of the Female Reproductive System of Abedus ovatus Stal (Belostomatidae: Hemiptera: Insecta) T G Lalitha+, K Shyamasundari, K Hanumantha Rao Department of Zoology, Andhra University, Visakhapatnam 530 003, A.P., India +Research fellow of University Grants Commission, New Delhi, and corresponding author. Fax: +91-891 555547
Received 13 November 1995
Code Number:OC97028
Sizes of Files:
Text: 21.1K
Graphics: Line drawings (gif) - 44.5K
Photographs (jpg) - 191.0K (3 files)
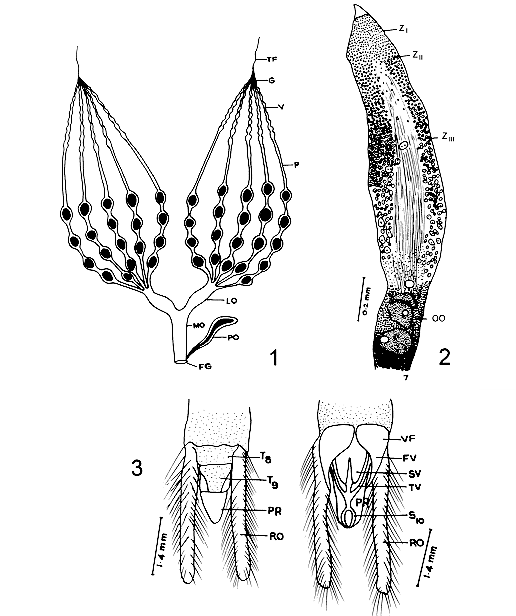
In Abedus ovatus Stal, the female reproductive system consists of a pair of ovaries leading to oviduct and opens out by genital aperture. Each ovary is composed of five telotrophic ovarioles, with four different regions, terminal filament, germarium, vitellarium and pedicel. Germarium and vitellarium are the germ tubes in which the development of oocyte occurs. In the germarium, the differentiation of trophocytes into young oocytes was observed in three zones, ZI, ZII and ZIII. Further development of oocytes in the vitellarium could be divided into seven stages, STI STVII. The developed eggs are lodged in the pedicel. Key words: ovary germarium vitellarium oocytes Morphological and histological studies on reproductive system of insects have called the attention of several investigators (Snodgrass 1935, Bonhag 1958, Davey 1965, Wigglesworth 1965). Information on the oogenesis of Popilus disjunctus Illiger, Laccotrephes maculates Fahr. Oncopeltus fasciatus Dallas, Schistocerca gregaria Forskol, Dytiscus, Cybister and Hygrobia and Triatoma infestana Klug were given by Bryan (1954), Bonhag (1958), Nath et al. (1959), Lusis (1963), Urbani (1970) and Barth (1973) respectively. Choi Won Chul and Walter (1976) made electron microscopic studies on the differentiation and development of trophocytes and oocytes in Gerris najas De Geer. Later the histological details of the ovarian structure is given by Deshpande and Srivastava (1981) in Dysdercus Koenigii Fabr and Ma and Ramaswamy in Lygus lineolaris. Hamilton (1931) and Kaushik (1972) contributed information on genitalia of Nepa cinerea Linn and Belostoma indicum Lep and Serv. Abedus ovatus is a freshwater bug with slightly greenish hue and transluscent body measures: 2025 mm in length. They commonly lurks among the phytal fauna in pools and ponds and its favourate resting position is to cling to some support with the tip of the abdomen. The most interesting and characteristic feature of this bug is that the female species lay their eggs on the back of the males and these encumbered males carry their eggs till they hatch. Females swimming in a rather ungainly manner and males with eggs on the back have called our attention to study the morphological and histological studies of the female reproductive system. MATERIALS AND METHODS Adult specimens were collected from Kondakarla Lake, 40 km from Visakhapatnam and maintained in the aquarium tanks in the laboratory. Visakhapatnam is located on the east coast of India between latitude 17 degrees 42' N and longitude 82 degrees 18' E. Ten female specimens were dissected for reproductive system and the material was fixed immediately in Susa or Zenker or 10% formalin or formol calcium. After processing 8 mm thick sections were cut in sagittal and transverse planes and the sections were stained with Azan and Mallory triple stain (Barth 1953, Pearse 1968) techniques for their histological study. The study material is deposited in the Museum, Zoology Department, Andhra University, Visakhapatnam, Register No.1979. RESULTS The female reproductive system of A. ovatus consists of a pair of ovaries which lie ventrolaterally in the body cavity of the abdomen on either side of the alimentary canal. Each ovary is composed of five telotrophic ovarioles, which are attached to the dorsal wall of the anterior prothorax by apical filaments. Posteriorly the ovarioles join to form a short lateral oviduct which inturn leads into a central median oviduct and opens out by female genital opening. The median oviduct receives a duct from paragenital organ (Fig. 1).
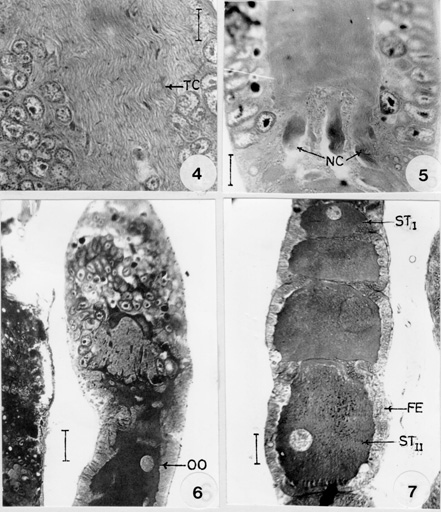
Fig. 2 : longitudinalsection of the germarium. ZI: zone-I. ZII: zone-II. ZIII: zone-III. OO: young oocytes. Fig. 3: genitalia (dorsal and ventral views). FV: first valvulae. PR: proctiger. RO: retractile organ. SV: second valvulae. S10: sternal segment. TV: third valvulae. T8, T9: tergal segments. VF: first valvifer. Each ovariole is composed of four main regions: (1) the threadlike apical filament, (2) the germarium, a chamber housing anteriorly the nurse cells and posteriorly the young oocytes, (3) a long vitellarium composed of a series of oocytes in their follicular sheaths, which become progressively large towards the posterior end, and (4) the pedicel in which the mature eggs are lodged before passing into the lateral oviduct. Genitalia is formed by the modification of eighth and ninth abdominal segments. The ovipositor is constituted by three pairs of compressed platelike valvulae and one pair of valvifer, each articulating with a small basal sclerite on the eighth and ninth tergum with lateral retractile organ and a central protiger covered with tenth sternum ( Fig. 3). The germarium is the most anterior chamber of the ovariole and it contains trophocytes, young oocytes and prefollicular tissue. A large portion of the anterior of the germarium is occupied by trophocytes, the apical position of the trophocytes is typical of the Hemiptera and for this reason the ovarioles of these insects are termed telotrophic ovarioles. In A. ovatus three zones, zone-I, zone-II and zone-III could be recognized in the trophic region (Fig. 2). Zone-I is the most anterior region and is characterized by small cells with scanty cytoplasm and spherical nuclei. Mitotic figures are found anteriorly in this zone, indicating the site for the origin of new trophocytes. Some of the nuclei have conspicuous and peripherally arranged condensed chromatin bodies. Zone-II is distinguished by its large cells which are three or four times the diameter of zone-I cells. They are irregular in shape. The nuclei are ovoid or angular and contain condensed chromatin bodies. Zone-III occupies more than two thirds of the trophocyte region and contains fully differentiated trophic tissue. This zone is characterized by the presence of branching strands of cytoplasmic material which eventually converge centrally and posteriorly to form an elongate trophic core. These strands are often extensions of intact cells and they apparently serve as pathways for the flow of material from degenerating cells into the trophic core. In the longitudinal section of the trophic core, longitudinal striations were observed throughout its length suggesting a fibrillar or lamellar composition (Fig. 4).
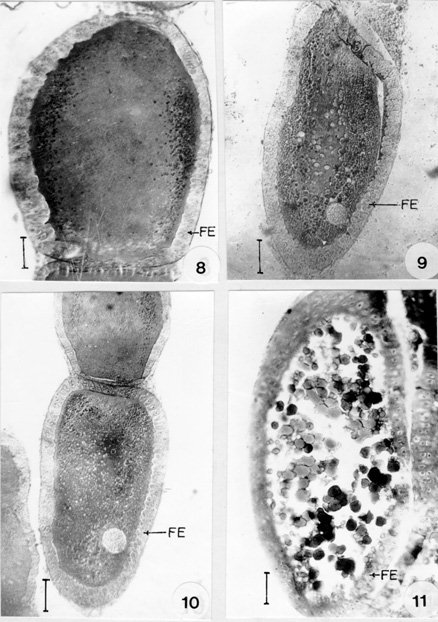
Fig. 5: posterior region of the germarium showing the presence of nutritive cords (NC). Bar 15 mm. Fig. 6: longititudinal section of the ovariole showing the formation of young oocytes (OO). Bar 15 mm. Fig. 7: longitudinal section of vitellarium showing stage-I (STI) and stage-II (STII) oocytes. FE: follicular epithelium. Bar 25 mm. In the anterior region of this zone, the cells are larger than those of zone-II. The size of the cells increases towards the posterior region and in their greatest dimensions they average approximately five times the diameter of zone-I cells. Their nuclei are similarly enlarged with rod or oval shape. Binucleate cells are found in this zone. From the posterior end of the trophic core, the nutritive cords radiates and join the prefollicular chamber (Fig. 5). These cords originate as small connecting strands between the base of the core and the youngest oocytes which are situated at the anterior border of the prefollicular region. Larger cords are connected with the most posterior oocytes and maintain their continuity with the oocytes as they move posteriorly into the vitellarium. Like the trophic core its branches are longitudinally striated. In the most posterior region of the germarium, the oocytes are small and spherical with large amounts of cytoplasm with a single nucleus (Fig. 6). The ooplasm of the young oocytes is homogenously granular with a centrally located nuclei. This condition persists throughout the stages, where the oocytes are surrounded by two or three layered epithelium. During the previtelline period the oocyte grows and elongates. Yolk has been observed peripherally in the oocytes as small globular spheres. In the young oocytes the nucleus is spherical and centrally located. As the oocyte further enlarge, the nuclei develop into germinal vesicle and increases in size due to the accumulation of vesicular fluid. The oocytes development in the vitellarium could be divided into seven stages (STI-STVII). In stage-I (Fig. 7) the follicular epithelium is multilayered and undergoes rapid division of growth. The oocytes measure up to 46.3 x 2.1 mm in diameter. The ooplasm is finely granular and its nucleus is spherical and more or less located in the centre.
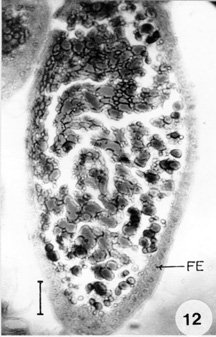
Fig. 9: stage-V oocyte. Bar 35 mm. Fig. 10: stage-IV oocyte. Bar 60 mm. Fig. 11: stage-VI oocyte. Bar 60 mm. FE: follicular epithelium. In stage-II (Fig. 7) the follicular epithelium is double layered in this stage. The oocytes measure 101.4 x 66.3 mm in diameter with a finely granulated cytoplasm. The seminal vesicle increases in size which is situated towards the periphery. In stage-III (Fig. 8) the follicular cells are arranged in a single row. The ooplasm is finely granular with peripherally placed yolk granules, which are seen as small spherical spheres. The oocyte measures up to 202.8 x 128.7 mm in diameter. Transparent spheres are sometimes observed in the region where the nutritive cords enter the oocytes. In stage-IV (Fig. 10) the follicular cells become cuboidal at this stage. The oocyte measures 269.1 x 206.7 mm in diameter. Large and small dense droplets and spheres are seen in the peripheral ooplasm. In stage-V (Fig. 9) binucleate columnar cells are observed in the epithelium. The oocyte measures up to 400 x 160 mm in diameter. The yolk droplets and spheres increases in size and number. Small droplets are oriented towards the periphery of the ooplasm. In stage-VI (Fig. 11) the follicle cells are squamous and binucleate. The oocyte is ellipsoidal in shape, measures about 608 x 224 mm in diameter. The ooplasm is completely filled with yolk droplets.
In stage-VII (Fig. 12) the sqamous cells of the epithelium become vacuolated and begin to breakdown. Formation of chorion layer is completed by the end of the vitellogenesis. The oocyte is packed with large yolk spheres and measures 672 x 320 mm in diameter. It shows tremendous increase in size before vitellogenesis. The follicular plug develops as a mass of cells in the developing oocytes of the vitellarium. As the oocytes development progresses the follicular cells extend spread all around the oocytes, thus separating the two successive oocytes. The pedicel is a simple tube, which lodges the developed eggs and connects vitellarium and the lateral oviduct. The wall of the pedicel shows the presence of squamous cells whose nuclei are ovoid and small in comparison with those of the follicular epithelium and contain homogeneously distributed chromatin granules. DISCUSSION In A. ovatus, the female reproductive system consists of a pair of ovaries situated on the ventral side of the abdomen on either side of the alimentary canal. Each ovariole is composed of five telotrophic ovarioles, leading to lateral oviduct which unite posteriorly to form a common oviduct. Anteriorly five ovarioles are joined by terminal filament by which they are attached to the abdomen. Each ovariole is composed of a slender terminal filament, germarium, vitellarium and pedicel. The germarium and vitellarium are the germ tubes in which the development of the egg takes place. The developed eggs are lodged in pedicel. The general organization is in agreement with the reproductive system of other hemipterans like Oncopeltus fasciatus (Bonhag & Wick 1953), Rhodnius prolixus Stal (Vanderberg 1963), Gerris regimis Say (Eschenberg & Dunlap 1966) and Dysdercus fasciatus Gign (Brunt 1971). Ma and Ramaswamy (1987) who followed the histological changes during first gonotrophic cycle, states that there are three distinct stages, previtellogenic, vitellogenic and choreogenic. Sareen et al. (1990) could make out six zones (Z1Z6) including the terminal filament in Lepisma saccharina L. The general morphology of the female genitalia in A. ovatus is in agreement with the description of the genitalia of other hemipterans (Hamilton 1931, Kaushik 1972). Mitotic figures have been observed in the anterior zone of the germarium of the ovariole of A. ovatus, indicating as it is the site of the origin of new trophocytes. Same reports were encountered in O. fasciatus by Bonhag (1958), R. prolixus by Vanderberg (1963) and Gerris remigis by Eschenberg and Dunlap (1966) but however, this report is in contrast to that observed by Davis (1956) in Cimex lectularis Linn. In most of the insects germarium is very active in the production of nurse cells a phenomenon which is a continuous process. In the germarium of A. ovatus the presence of cytoplasmic units with one or two prominent nuclei were observed as reported by Das Gupta and Ray (1954) in Cimex. However, syncytial character of this region has been observed by Schrader and Leutchenberger (1952) in Acanthocephala and by Bonhag and Wick (1953) in O. fasciatus. Longitudinal striations were observed in trophic core of the germarium of A. ovatus. The striated conditions of the trophic core is reported in Adelphocoris lineolatus Goeze (Masner 1966) and in D. fasciatus (Brunt 1971). In A. ovatus, the nutritive cords were connected to the developing oocytes, which are originated from trophic core. Nutritive cords development was reported in other insects like Popilius disjunctus (Bryan 1971). In A. ovatus, the prefollicular tissue migrates posteriorly into multilayered columnar epithelium which surrounds young oocytes and later give rise to the follicular epithelial cells. Brunt (1971) reported the same findings in D. fasciatus. Binucleate condition is observed in the follicular epithelial cells in the vitellarium of A. ovatus as it is reported in O. fasciatus (Bonhag & Wick 1953). ACKNOWLEDGEMENTS To Andhra University authorities for providing facilities to work. REFERENCES
The following images related to this document are available:Photo images[oc97028b.jpg] [oc97028f12.jpg] [oc97028f1-3.jpg] [oc97028d.jpg] [oc97028f8-11.jpg] [oc97028f4-7.jpg] [oc97028c.jpg]Line drawing images[oc97028a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}