 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Age structure of adult mosquito (Diptera: Culicidae) populations from Buenos Aires Province, Argentina Arnaldo Macia Centro de Estudios Parasitológicos y de Vectores (CEPAVE), Calle 2 No. 584, 1900 La Plata, Prov. de Buenos Aires, Argentina This work is part of author's doctoral Thesis, Facultad de Ciencias
Naturales y Museo, Universidad Nacional de La Plata.
Received 7 May 1996
Code Number:OC97032
Sizes of Files:
Text: 29.6K
Graphics: Line drawings and tables (gif) - 111K
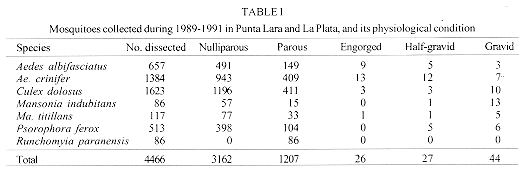
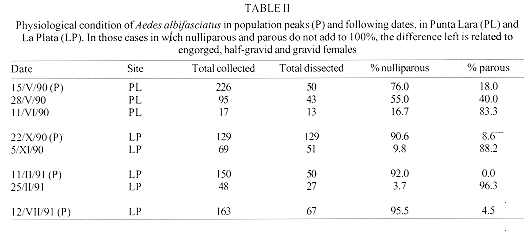
In order to detect seasonal trends in the age structure of adult mosquitoes from Buenos Aires province, Argentina, female populations were sampled with CDC traps during 1989-1991 in Punta Lara and La Plata. The mosquitoes were dissected and age-grouped according to ovarian tracheation and ovariolar stages. All Runchomyia paranensis females were parous, suggesting that this species could be autogenous. Aedes albifasciatus showed parous peaks following population peaks, with shorter delays in spring-summmer and longer in fall-winter. Ae. crinifer and Culex dolosus showed wide fluctuations in age structure due to adult emergences during all months. Psorophora ferox showed high population replacement rates. Mansonia indubitans and Ma. titillans have few generations per year during their activity period. This is the first report on age-grading of adults of field mosquito populations from Argentina. Key words: mosquitoes - adults - age structure - Argentina Age structure of adult mosquitoes is important from the ecological and epidemiological points of view. Age structure studies could provide estimates of survivorship and longevity and the likelihood of infection for non-transovarially acquired pathogens. Age determination could be used to evaluate control measures success against the vector population. Detinova (1962) and Service (1993) reviewed the age-grading methods available for adult culicids. Reproductive categories of female mosquitoes give information on temporal changes in the age structure of populations. Based on this feature, several authors have shown the temporal pattern of adult population abundance in species belonging to Aedes (Packer & Corbet 1989, Sokolova 1994), Anopheles (Gillies & Wilkes 1965, Charlwood & Wilkes 1979, Jensen et al. 1993), Culex (Bellamy & Reeves 1963, Burdick & Kardos 1963, Nelson 1971, Mitchell 1979, 1981, 1983, Reisen et al. 1983, 1986) and Mansonia (Corbet 1963, Service 1969, Lounibos et al. 1990). Previous papers (Ronderos et al. 1992, Campos et al. 1995, García et al. 1995, Macia et al. 1995, 1996) reported seasonal changes in the abundance of Ae. (Ochlerotatus) albifasciatus (Macquart), Ae. (Ochlerotatus) crinifer (Theobald), Cx. (Culex) dolosus (Lynch Arribalzaga), Mansonia (Mansonia) indubitans Dyar and Shannon, Ma. (Mansonia) titillans (Walker), Psorophora (Janthinosoma) ferox (Humboldt) and Runchomyia (Runchomyia) paranensis (Brethes), the main species in a taxocenosis from Buenos Aires province. Ae. albifasciatus, Ae. crinifer and Ps. ferox are floodwater mosquitoes. Immatures of Ae. albifasciatus are found mainly in grassy, sunlit, shallow depressions of ground flooded by heavy rains. When these depressions are flooded after a long period of dryness, breeding places result in explosive increases of populations. These high levels occur during spring and fall, the species being scarce for most part of the year (Luduena Almeida & Gorla 1995, Macia et al. 1995). Typical breeding places for Ae. crinifer are similar, but in shaded places under the tree canopy. Larvae are found in any month, peaking during spring and fall. Populations do not show explosive increases (Macia et al. 1995). Larvae and pupae of Ps. ferox inhabit shaded ground pools in forests. Population density fluctuates according to water level of habitats and environmental temperatures, and the immatures occur between September and April (Campos et al. 1995). Immature stages of Cx. dolosus, Ma. indubitans and Ma. titillans develop in permanent water bodies. Cx. dolosus breeds in natural and artificial habitats, mainly in permanent ponds with aquatic vegetation and stagnant water. Immatures can also be found in a great variety of small to large places with clear to turbid water, in stream margins, and other places (Almirón & Brewer 1996). Immatures of both species of Mansonia could be found in water bodies covered with floating vegetation. Larvae and pupae attach to roots and leaves of water lettuce, Pistia stratiotes L. Seasonality of Mansonia is associated to phenology of host plants at the latitude of Buenos Aires province (Garcia et al. 1995). Larvae of Ru. paranensis remains unknown (Darsie & Mitchell 1985). This work continues the above mentioned paper series, and reports on the seasonal trends in variations of adult age structure for these taxa, and their relationship with population fluctuations. MATERIALS AND METHODS The study was carried out at two sites in Buenos Aires province. The first, Punta Lara, is located near Rio de La Plata (34 degrees 51'53"S, 57 degrees 52'23"W). The site is included in a zoogeographic ecotone between the Pampasic and Subtropical domains (Ringuelet 1981), altered by man. Mosquito collections were made inside a gallery forest where the main species are "ligustro" (Ligustrum lucidum Ait.), "seibo" (Erythrina cristagalli L.), "cortadera" (Scirpus giganteus Kunth) and "zarzamora" (Rubus ulmifolius Schott). The terrain is frequently flooded by tides from Rio de la Plata estuary, rains and overflow of streams. The other site, 15 km away, is a field in the urban zone of La Plata city, belonging to the School of Astronomic Sciences, National University of La Plata. Mosquito collections were performed in a field with scattered trees. There is a stronger influence of winds and sunlight than in Punta Lara due to the absence of high vegetation. Shallow water bodies appear in land depressions after rains. In both places, host-seeking females were caught fortnightly during a 24 hr period, using two CDC traps supplied with ligth and CO2 (Service 1993). Traps of the same sampling station were 15 m distant. In Punta Lara, 42 samples were obtained between August 1989 and July 1991. In La Plata, only 27 catches were performed between August 1990 and July 1991. Maximun and minimun daily temperatures were obtained for the whole period. The mosquitoes were sorted to species according to Ronderos and Bachmann (1963) and Darsie and Mitchell (1985). Mosquitoes collected by one of the traps were used to obtain data about female relative abundance and population levels. All mosquitoes collected by the other trap were frozen (-18 C) to arrest ovarian development and used to establish the physiological status of females. A minimun of 10% of frozen samples, or the whole sample when it contained less than 50 females, were dissected following the technique described in Garcia et al. (1994). Females were scored according to their ovarian development stages (Christophers 1911). Empty females (with follicles in stage I or early II) were classified as either nulliparous or parous by the tracheolar skeining technique (Detinova 1962). Females with ovaries reaching stages II to V were classified into the following categories: engorged: midgut with fresh blood, follicles in stage II or III; half-gravid: ovaries extending over 4-5 abdominal segments in ventral view and 6 in dorsal view; follicles in stage III or IV; gravid: follicles in stage V; empty: midgut without blood, follicles in stage I or II. The total annual percentage of parous females was computed by multiplying by 100 the quotient between the sum of parous, engorged, half-gravid and gravid females, and the number dissected. For this purpose, it was assumed that every engorged, half gravid and gravid females would complete the gonotrophic cycle, so their numbers were added to that of the parous. Monthly parous rates were computed as the quotient between number of parous females and total number of females caught, then multiplying it by 100. RESULTS AND DISCUSSION Ae. crinifer, Ps. ferox and Ru. paranensis were collected only in Punta Lara. Data on the remaining species, collected in both sites, were considered as belonging to only one population. A total amount of 4,466 mosquitoes from Punta Lara and La Plata were dissected. About 71% were nulliparous, 27% were parous, and the rest engorged, half-gravid, and gravid (Table I). Most species were represented mainly by young females. However, this was not the case in Ru. paranensis, because all of the 86 dissected individuals (1989-1990 in Punta Lara) were parous. All females of this species were in Christophers's early stage II, suggesting that females complete their first oviposition autogenously. If Ru. paranensis were autogenous, the mosquitoes could be attracted by hosts only after completion of the first gonotrophic cycle. Thus, CDC traps would catch only parous individuals. This hypothesis still needs confirmation. In the other species, the total annual percentage of parous females was high (>30%) in Ae. crinifer (31.9%), Ma. titillans (34.2%) and Ma. indubitans (33.7%), and low (<30%) in Cx. dolosus (26.3%), Ae. albifasciatus (25.3%) and Ps. ferox (22.4%). These percentages are the estimated population that seeks more than one blood meal. Thirty per cent or more was considered high by Morris and DeFoliart (1971), and it indicates, a priori, species with higher field survivorship than those with lower percentages.
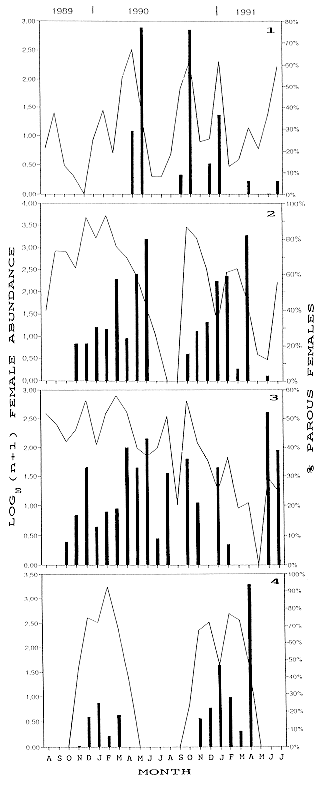
Figs 1-4: monthly total fluctuation of number of mosquitoes collected in Punta Lara and La Plata, 1989-1991 (solid line) and percentage of parous females (columns). 1: Aedes albifasciatus.2: Aedes crinifer. 3: Culex dolosus. 4: Psorophora ferox Monthly percentages of Ae. albifasciatus parous females ranged from 0% in June 1991 (all nulliparous females) to 77% in June 1990. A second peak of 76% was detected in November 1990 (Fig. 1). The number of parous females peaked during the sampling dates following Ae. albifasciatus population peaks (Table II). This succession of maximum levels of population and maximun levels of parous individuals was not observed in the other species. The population peaks of Ae. albifasciatus comprised high percentages of nulliparous, indicating high recruitment dates. On the following dates, the diminishing overall number collected coincided with increased parous rates and a decrease in nulliparous, suggesting a reduction in the number of emerging adults from breeding sites. Each Ae. albifasciatus population peak was related to a parous peak recorded 15 or 30 days later, depending upon the season of the year. Thus, the maximum parous rate recorded on 11 June 1990 (83.3%) in Punta Lara followed after about 30 days the population peak recorded on 15 May 1990. The average temperature between both dates was 12.2oC. On the other hand, there was a delay of only one sampling date (about 15 days) among parous peak corresponding to 5 November 1990 and the population peak of 22 October 1990 in La Plata, when the average temperature of the period was 18 C. A similar situation was observed between 11-25 February 1991, in La Plata, when the average temperature was 20 C. Lower temperatures in May and June, and higher temperatures in October, November and February could account for those differences in delays between peaks. Therefore, this shows that the time elapsed between the emergence of an adult brood and their first oviposition is shorter than 15 days in spring-summer and about one month in fall-winter. In laboratory studies, Ludueña Almeida and Gorla (1995) found that most of females kept under controlled conditions laid eggs 5-8 days after a blood meal, although some oviposited after 19 days. The difference of these results compared to the ones of the present study, could be due to the time elapsed between emergence and blood feeding. These authors reported an average of 3.8 gonotrophic cycles (maximun 5) for Ae. albifasciatus field-collected females, based on the method of follicular relics (Ludueña Almeida & Gorla 1995). Monthly parous rate of Ae. crinifer ranged from 3 to 82% (June and April 1991, respectively) (Fig. 2). There was another peak (80%) in the parity rate during June 1990. There was no clear relationship among population levels and parous rates. The latter tended to increase as the number of females decreased during the cooler months. Monthly parous rates of Cx. dolosus ranged from 7 to 52% (February and June 1991, respectively) (Fig. 3). It is difficult to generalize trends about parous rates related to seasonal changes in female abundance. Adults of Ae. crinifer and Cx. dolosus can be found throughout the year. Both species are multivoltine (Macia et al. 1995 for Ae. crinifer, Macia et al. 1996 for Cx. dolosus). There was wide variation in population numbers during short periods, and a continuous output of nulliparous females during all months. These facts masked trends in age-structure undergoing deep fluctuations. This irregularity is a feature of mosquito dynamics from moderate latitudes, due to the changing environmental conditions, which determine intensity of emergences and mortality in each physiological age (Detinova 1968). Monthly parous rates of Ps. ferox were low, as compared with other species. Host-seeking females occur from October to May (Fig. 4). During November 1989 all females were nulliparous just emerged from breeding places. The highest percentage of parous individuals was recorded during April 1991, coinciding with a population decrease. This agrees with data shown by Morris and DeFoliart (1971). These authors observed in Ps. ferox from USA that almost all females become parous when population declines. In Punta Lara, during the first sampling year (1989-1990), 14.6% (27/184) of females were parous. In 1990-1991, parous females increased to 23.4% (77/329). The lower parous rates detected in 1989-1990, as compared to 1989-1990, could have occurred as a consequence of a sampling artifact. A fast development is typical of this species (Forattini 1965), and sampling frequency in this study could exceed immature development time and gonotrophic cycle lenght. Monthly parous rates reported here could be lower than real values, if peaks in parous rates were not well represented in the samples. As pointed above, Ps. ferox had the lowest total annual proportion (22.4%) of parous females of the species analized herein, suggesting low survivorship in nature. In contrast, Morris and DeFoliart (1971) found that Ps. ferox had the highest parous rate among several species of Aedes, Culex, Culiseta and Mansonia from USA, and they concluded that Ps. ferox had a high survival rate, one out of two females living enough to seek a second blood meal. In the present study, low percentages of parous mosquitoes could be a consequence of large increases in emergence due to many floodings in larval habitats during 1989-1991 (Campos et al. 1995). Ma. indubitans was collected from October to May in Punta Lara, and from January to April in La Plata. Almost all females collected in March-April 1990 from Punta Lara were nulliparous, and a few parous individuals were collected nearly at the end of that period. During January 1991, both in Punta Lara and La Plata, most females were gravid. Around 70% of females were nulliparous during February and March. Parity rate ranged from 5 to 50% from mid-February to April (Fig. 5).
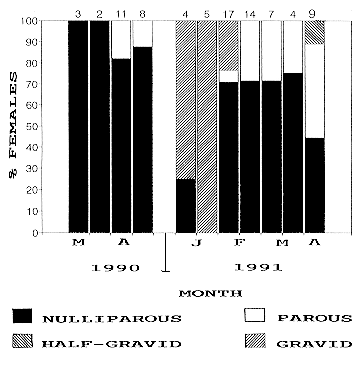
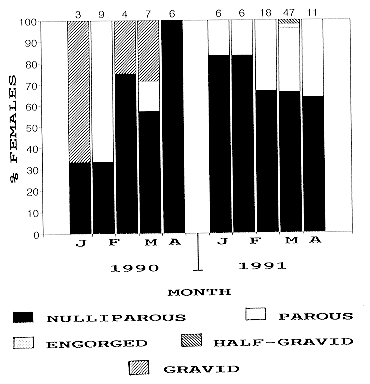
Fig. 5: variation in percentages of nulliparous, parous, half-gravid and gravid Mansonia indubitans females from Punta Lara and La Plata, 1990-1991. Numbers on columns indicates number of individuals dissected in the corresponding period. Engorged females not found in this species. Females of Ma. titillans exhibited host-seeking activity from December 1989 to May 1990, and from October 1991 to May 1991 in Punta Lara. The pattern was somewhat different in La Plata, where the species was collected from January to April. During 1990, Ma. titillans gravid females were present from early January to early March. In April, the whole population consisted of nulliparous females. During 1991, parous individuals ranged from 16 to 60% from January to May (Fig. 6).
Fig. 6: variation in percentages of parous, engorged, half-gravid and gravid Mansonia titillans females from Punta Lara and La Plata, 1990-1991. Numbers on columns indicates number of individuals dissected in the corresponding period. The age structure data for Ma. indubitans and Ma. titillans seems to reflect the existence of few adult generations per year with slight overlap between them. Population levels for Mansonia mosquitoes were low. Ma. indubitans and Ma. titillans have three or four annual generations near the north of their distribution (Provost 1976). In Punta Lara and La Plata, this could be similar. If these species overwinter in the adult stage in the study area (Garcia et al. 1995), overwintered females could start host-seeking activity in October, when first individuals were caught by the CDC traps. These overwintered females then laid eggs, and gave origin to the larval population. Larvae appear in breeding places in December (Ma. titillans) or January (Ma. indubitans) (Garcia et al. 1995). High numbers of gravid females collected during January, simultaneously with immature development, may reflect increased reproductive activity during warmer months. Towards fall, increased parous rates could be due to increased number of "older" females at the end of the annual cycle. In every species mentioned in the present study, nulliparous and parous females had ovarian follicles in early stage II. Engorged females had follicles in late stage II or early III; half-gravid females, in late III or IV; and gravid females, in stage V. Therefore, all the mosquitoes were in gonoactive phase of the gonotrophic cycle, and there was gonotrophic concordance (Clements 1992) in the species studied. Several reserchers have studied daily survival rates based on parity rates of mosquito populations (Davidson 1954, Gad et al. 1989, Gillies & Wilkes 1963, Weathersbee & Meisch 1991). These estimates cannot be apllied to this study because none of the populations from Punta Lara and La Plata was found to be in steady-state age distribution in any period, a condition that must be fulfilled to obtain such estimates (Service 1993). The parous rates underwent frequent wide fluctuations during the sampling period. In all species from both sites, recruitments and losses were not equal. As a second reason to invalidate survival estimates, the length of the gonotrophic cycle in nature is unknown for the species treated herein. The trapping method used in this study could have affected the parous rate estimates. The sampling procedure based on biweekly collections may have not been adequate enough to describe the age structure of populations from Punta Lara. During the field work it was observed that breeding places flooded and dried several times between two consecutive sampling dates, sometimes more than once a week. Traps were located close to ephemeral pools, thus increasing the probability of collecting younger females. Therefore, parous rates reported in this paper may be biased and should be considered as an underestimation of real values that occur in nature. ACKNOWLEDGEMENTS To RE Campos for help during field works. To the personnel of School of Astronomic and Geophysical Sciences, National University of La Plata, for providing climatic data. REFERENCES ME 1983. Population ecology of Culex tarsalis (Diptera: Culicidae) in a foothill environment of Kern County, California: Temporal changes in female relative abundance, reproductive status, and survivorship. Ann Entomol Soc Am 76: 800-808. Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Line drawing images[oc97032d.gif] [oc97032c.gif] [oc97032e.gif] [oc97032b.gif] [oc97032a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}