 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biomorphological alterations induced by an anti-juvenile hormonal compound, 2-(2-ethoxyethoxy)ethyl furfuryl ether, on three species of Triatominae larvae (Hemiptera, Reduviidae) Jose Jurberg, Cleber Galvao/+, William S Bowers*, Eloi S Garcia**, Patricia Azambuja** Laboratorio Nacional e Internacional de Referencia em Taxonomia de
Triatomineos, Departamento de Entomologia +Corresponding author. Fax: +55-21-290.9339. e-mail: galvao@dcc001.cict.fiocruz.br Part of this paper was presented in the XXI Annual Meeting on Basic Research in Chagas'Disease, Caxambu, MG, Brasil, 1994. Received 31 October 1996
Code Number:OC97052
Sizes of Files:
Text: 17.4K
Graphics: Line drawings and tables (gif) - 33.5K
Photographs (jpg) - 55.4K
Applied topically to larvae of Rhodnius prolixus Stal, Triatoma infestans (Klug) and Panstrongylus herreri Wygodzinsky, vectors of Trypanosoma cruzi, the causative agent of Chagas'disease, a synthetic, furan-containing anti-juvenile hormonal compound, 2-(2-ethoxyethoxy)ethyl furfuryl ether induced a variety of biomorphological alterations, including precocious metamorphosis into small adultoids with adult abdominal cuticle, ocelli, as well as rudimentary adultoid wings. Some adultoids died during ecdysis and were confined within the old cuticle. The extension of these biomorphological responses is discussed in terms of the complexity of the action of anti-juvenile hormonal compounds during the development of triatomines. Key words: Rhodnius prolixus - Triatoma infestans - Panstrongylus herreri - adultoid - anti-juveline hormone From the beginning of the studies of the action of precocenes in insects (Bowers 1976, 1977, Bowers et al. 1976), the compounds with anti-juvenile hormonal (AJH) activity are considered important new approaches to insect control. These compounds cause the developing of premature stages, i.e., they induced diminutive and sterile adults named adultoids (Bowers 1976, 1977, 1982, Azambuja et al. 1981). Phytochemicals with AJH activities in insects have been discovered in several plants (Bowers et al.1976, Garcia et al. 1984, Bowers & Areguilin 1987, Garcia & Azambuja 1987). Several AJHs have been obtained by synthetic chemical routes (Quistad et al. 1981, Kuwano et al. 1983, Staal 1986, Brooks et al. 1988). The effects of precocenes were investigated in different triatomine species. For example, Azambuja et al. (1981), Garcia et al. (1984, 1988) and Jurberg et al. (1982, 1984, 1986) described the actions of precocene in Rhodnius prolixus, Panstrongylus megistus and Triatoma infestans. Recently, it has been demonstraded that novel furanyl ethers have potent AJH activity in Oncopeltus fasciatus Dallas (Bowers et al. 1995) and in triatomines (Azambuja et al. 1996) inducing precocious metamorphosis and other modifications in these insects. In this investigation we examine the biomorphological effects of 2-(2-ethoxyethoxy)ethyl furfuryl ether (EEFE), one of the most active of the furanyl ether AJHs, on the hematophagous bugs, R. prolixus, T. infestans and Panstrongylus herreri, three vectors of Trypanosoma cruzi, a protozoan that causes Chagas'disease in Latin-America. MATERIALS AND METHODS Insects and feeding - Carefully staged third-instar larvae of R. prolixus Stal, 1859, T. infestans (Klug, 1834) and P. herreri Wygodzinsky, 1948, reared and maintained in the Department of Entomology, FIOCRUZ, were used throughout this investigation. Weights just before feeding ranged from 4.5 mg for Rhodnius to 7.6 mg for Pastrongylus. Following ecdysis, the insects were starved for 30-35 days and then fed on mouse. Only fully engorged insects were used in all treatment. The insects were maintained at 28oC during the whole experiment as described previously (Garcia et al. 1988). Topical treatment - Topical treatments were performed on groups of 20-30 third-instar larvae of these triatominae species. EEFE at a concentration of 60ug in 0.5ul of acetone was applied to the ventral surface of the abdomen on the fourth day before feeding. The biomorphological alterations induced by EEFE were determined by the external inspection of the adultoids produced, in particular, on the external genitalia of males of these three species of triatomines. Controls received solvent treatment only. Genitalia procedures - Genitalia was taken up from insects with morphological alterations, heated in 10% KOH, and manipulated in phenol up to the mounting in Canada balsam as previously described by Jurberg et al. (1984). The figures were drawn using stereoscopic microscope connected to a camara lucida. RESULTS General external morphology - Visual inspection of the adultoids demonstrated that topical application with EEFE produced many R. prolixus adultoids with fully adult abdominal cuticle, ocelli, tibial adhesive organs, absence of the setiferous tubercle, pronotum within collar. Antero-lateral angles and anterior and posterior lobes were abnormal, and the rudimentary adultoid wings never attained the first abdominal segment but possessed full articulation with the thorax. General biomorphological alterations in the other two species of triatomines were similar to these described above for Rhodnius.
Figure 2a,b: induction of adultoids in the third-instar larvae of Triatoma infestans by the treatment of EEFE at a concentration of 60ug on the fourth day before feeding. Figure 2c: induction of adultoids in the third-instar larvae of Triatoma infestans by the treatment of EEFE at a concentration of 60ug on the fourth day before feeding.
We also observed that adultoids induced by topical application with EEFE had further morphological responses: (i) sometimes died during ecdysis and were confined within the old cuticle and, normally, the head was recurved to the ventral surface of the abdomen in all species tested ( Fig.1); these morphological alterations had different intensity varying with the period of moulting and the species of triatomines (Figs 2a,b,c and 3); (ii) in the adultoid of P. herreri the filiform antennae did insert far from the anterior to the eyes and (iii) R. prolixus presented pronotum and mesonotum separated, metanotum with the posterior border visible and rudimentary scutellum between the bases of the forewings; sometimes the scutellum was absent.
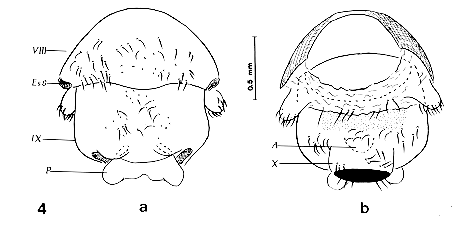
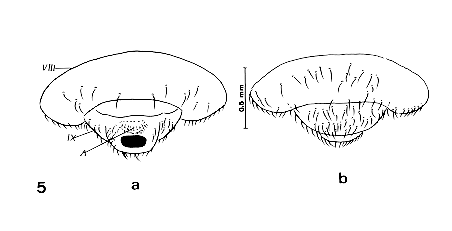
Figure 4: pygophore or 9th abdominal segment of an adultoid of Triatoma infestans obtained by the treatment of EEFE at a concentration of 60ug on the fourth day before feeding. a-ventral and b-dorsal views. VIII=8th segment; Es8=stigma of the 8th segment; IX=pygophore or 9th segment; P=phallus; A=anus; X=10th segment. Figure 5: last abdominal segments of an adultoid of Panstrongylus herreri obtained by the treatment of EEFE at a concentration of 60ug on the fourth day before feeding. a-dorsal and b-ventral views. VIII=8th segment; IX=pygophore or 9th segment; A=anus.
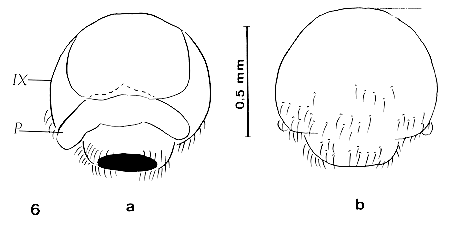
Figure 6: pygophore of an adultoid of Rhodnius
prolixus Male external genitalia - The main alterations of the external genitalia of males in adultoids of the three species of triatomines experienced are: (i) T. infestans presented abnormal phallus formed by a hard chitinized plate with any morphological characteristics related to normal structures including the absence of articulation that facilitates the phallus gets out of the pygophore (Fig. 4a,b); (ii) P. herreri had neither the male external genitalia nor the formation of globulous pygophore (Fig. 5); and (iii) R. prolixus adultoids had pygophore linked to the 10th abdominal segment. In addition, we observed both no genital structures inside the pygophore and an abnormal rectangular plaque with slightly curvature showing the base of the pygophore ( Fig. 6 a,b). The normal aspect of these structures is highly complex and therefore we prefer to refer to the figures already shown by Lent and Jurberg (1969,1975,1978). Finally, Table demonstrated by comparison that the morphological alterations induced by EEFE are similar to those induced by the classical AJH compound. DISCUSSION In the present paper, we investigated the biomorphological responses on larvae of three species of triatomines induced by the most active furanyl derivatives, EEFE. In fact, we already studied the effect of EEFE on several species of triatomines giving the paucity of information on the ecdysial stasis, delay of moulting, lethality, production of precocious adultoids under different modes of treatments, and comparison of this compound with precocene II (Azambuja et al. 1996). Now, we extend these findings and describe with more details the biomorphological alterations caused by the topical treatment of 3rd instar-larvae of R. prolixus, T. infestans and P. herreri. Our present results, considering in particular the male external genitalia, demonstrate considerable variation in sensitivity amongst species. For example, P. herreri was more sensitive to EEFE than R. prolixus and T. infestans if we observe the alterations induced in the male external genitalia. In the former species it was not found genitalia and formation of the characteristic pygophore, while the latter species demonstrated abnormal phallus (T. infestans) and pygophore (R. prolixus). However, these three species demonstrated about equivalent sensitivity when we consider the general external morphology. It is interesting to point out that the induction of precocious metamorphosis was greater in T. infestans than in R. prolixus and P. herreri (Azambuja et al. 1996). Since the mode of action, species specificity and spectrum of activity of the EEFE is presently unknow (Bowers et al. 1995, Azambuja et al. 1996) it seems valuable to speculate that (i) it is possible that the difference of sensitivity between the triatomine species studies was due to the dynamics of cuticular penetration, pharmacokinetics and metabolisms of the compound and (ii) EEFE may not affect the corpus allatum but terminate juvenile hormone (JH) production/release rather than alter or compete with JH cellular receptor, since precocious metamorphosis was inhibited by therapy with JHA (Bowers et al. 1995, Azambuja & Garcia unpublished data). Notwithstanding these speculations, our studies suggest that the furanyl-derivatives should be studied in other species of triatomines.
This work was supported by Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq) to ESG, PA and JJ, Programa de Apoio à Pesquisa Estrategica em Saúde (PAPES, FIOCRUZ) to PA, BIRD/FNS/FIOCRUZ contract number 027/93 and European Union (STD-TS3-CT-920092) to JJ and CG. REFERENCES and the reversal of ecdysial stasis by ecdysone. Arch Insect Biochem Physiol 1: 367-373. Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97052a.jpg] [oc97052b.jpg] [oc97052d.jpg] [oc97052c.jpg]Line drawing images[oc97052e.gif] [oc97052f.gif] [oc97052h.gif] [oc97052g.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}