 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
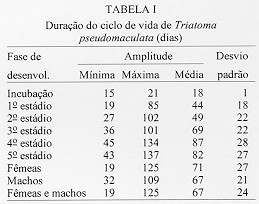
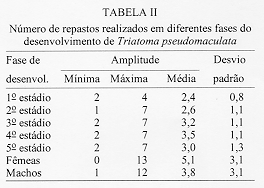
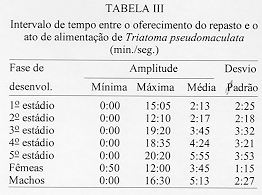
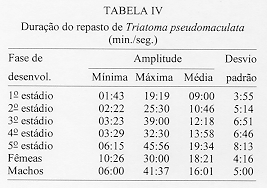
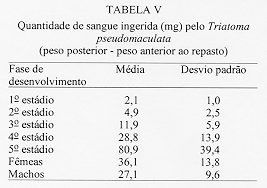
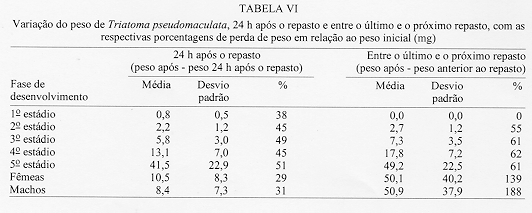
Alguns aspectos da biologia de Triatoma pseudomaculata Correa & Espinola, 1964, em condicoes de laboratorio (Hemiptera:Reduviidae:Triatominae) Teresa Cristina M Goncalves+, Vanda Cunha, Edson de Oliveira, Jose Jurberg Laboratorio Nacional e Internacional de Referencia em Taxonomia de Triatomineos, Departamento de Entomologia, Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil +Autor correspondente. Fax:& +55-21-590.9339.E-mail: Goncalves@DCC001.CICT. FIOCRUZ.BR Financiado pelo PAPES, FAPERJ, CNPq, Convenio 027/93 FNS/BIRD/FIOCRUZ Recebido em 11 de junho de 1996
Code Number:OC97054
Sizes of Files:
Text: 23.3K
Graphics: Tables (gif) - 208.4K
Biology of Triatoma pseudomaculata Correa & Espinola, 1964, under Laboratory Conditions (Hemiptera:Reduviidae:Triatominae) - Observations were made on the evolutive cycle of Triatoma pseudomaculata, held under laboratory conditions, fed weekly on bird (pigeon). Of 60 eggs obtained, only 34 nymphs reached the adult stage in a period of X(S)=398+/-76 days. The following parameters were observed: the time immature stages took to develop from egg to adult emergence; the occurrence of the first meal; the time-lapse between the presenting of the blood-meal and the begining of feeding; time of feeding; amount of blood ingested; variation of weight 24 hr after the blood-meal and until the next blood-meal; and the defecation pattern. The experiment was carried out for 20 months, held in BOD incubator with the average of temperature and humidity of 28+/-1 C and 80+/-5% RU, respectively. Key words: Triatoma pseudomaculata - biology - laboratory conditions Lucena (1959) refere-se a Triatoma maculata (Erichson, 1848), posteriormente Triatoma pseudomaculata Correa & Espinola, 1964, como uma especie de importancia regional, encontrada em pombais, galinheiros e cercas, sempre no peridomicilio e nunca em abrigos silvestres, e como caracteristica da fauna nordestina, distribuida geograficamente pelos Estados de Pernambuco, Paraiba, metade do Ceara, sertao de Alagoas, Bahia, Minas Gerais, Piaui e Goias. Silveira et al. (1984) relatam a ampla distribuicao geografica dessa especie, comentam sua baixa frequencia nas habitacoes, embora colonize algumas casas, e informam que, possivelmente, e o vetor do Trypanosoma cruzi em algumas areas da Paraiba. Dados da Fundacao Nacional de Saude, nao publicados, mostram que entre as sete especies presentes no Estado de Pernambuco, infestando o domicilio humano, ela ocupa o segundo lugar, porem com baixo indice de infeccao (dos 1.709 especimes capturados, 0,5% estava positivo). Este resultado pode estar de acordo com as observacoes de Perlowagora-Szumlewicz e Muller (1982) e Perlowagora-Szumlewicz et al. (1988) pois, segundo esses autores, a domiciliacao altera a relacao vetor-parasita reduzindo o indice de infeccao e a carga parasitaria. Dias (1991) comenta sobre a dificuldade da erradicacao dessa especie, uma vez que e nativa, ubiquista e ocorre em grandes quantidades no meio extradomiciliar, especialmente em regioes pobres e isoladas da area endemica. Forattini et al. (1981) classificaram-na como de moderada tendencia a domiciliacao mas com elevada ornitofilia, embora o estudo da preferencia alimentar tenha-lhe atribuido um ecletismo alimentar. Apesar de incluida entre as cinco especies mais importantes pela transmissao da doenca de Chagas no Brasil, foram poucos os estudos feitos a respeito de sua biologia (Heitzmann-Fontenelle 1972, Perlowagora-Szumlewicz 1976). Neste trabalho, procurou-se conhecer a biologia dessa especie utilizando como fonte alimentar a ave, tendo em vista, nao so a ornitofilia ja observada por Forattini et al. (1981), como tambem procurou-se evitar as modificacoes de parametros fisiologicos ocasionadas por alimentacao artificial (De Isola et al. 1980). Espera-se, com esses resultados, obter informacoes que contribuam para melhorar a compreensao da epidemiologia da doenca de Chagas nas areas de ocorrencia de T. pseudomaculata, e que facilitem o planejamento de acoes de controle, uma vez que a sua presenca no peridomicilio possibilitara constantes invasoes dos domicilios. MATERIAIS E METODOS O experimento iniciou-se com 60 ovos, que foram acondicionados em frascos, por postura; conforme ocorriam as eclosoes, as ninfas eram individualizadas em frascos de Borrel, devidamente identificados, contendo papel de filtro no fundo e dobrado em sanfona, para retirar o excesso de umidade proveniente das fezes e para aumentar a area livre. A alimentacao foi oferecida, semanalmente, com sangue de pombo (Columba livia). Para isso, depenava-se uma pequena area do flanco suficiente para a acomodacao do frasco de Borrel. Para as ninfas de 1 estadio o oferecimento foi diario, a fim de se observar a ocorrencia do primeiro repasto. Foram avaliados os seguintes parametros: periodo de incubacao; duracao de cada estadio ate a fase adulta; realizacao do primeiro repasto; intervalo de tempo entre o oferecimento e o inicio do repasto; duracao do repasto; quantidade de sangue ingerida; perda de peso ate 24 h apos o repasto e ate o proximo, em relacao ao peso inicial; numero de defecacoes por estadio, em um periodo de ate 10 min. apos a alimentacao. O experimento teve a duracao de 20 meses e foi realizado em estufa BOD a 28+/-1 C, 80+/-5% UR e fotoperiodo de 12 h. A pesagem foi realizada em balanca de precisao. O material trabalhado pertencia a uma colônia procedente de Morada Nova (CE) mantida no insetario do Laboratorio Nacional e Internacional de Referencia em Taxonomia de Triatomineos do Departamento de Entomologia do Instituto Oswaldo Cruz. RESULTADOS Dos 60 ovos obtidos, apenas 34 especimes (16 femeas e 18 machos) atingiram a fase adulta. Os demais (doze ninfas de 1o estadio, quatro de 2 e 4 estadios e tres de 3 e 5 estadios) morreram, na sua maioria, por nao conseguirem se alimentar, apesar de varias investidas sem sucesso, ou por motivo inexplicavel (tres casos). A duracao do ciclo do 1 estadio a adulto foi de X(S)=398+/-76 dias, com o minimo e maximo de 269 e 532 dias, respectivamente. Do 1 estadio ate a ecdise imaginal das femeas, o ciclo foi de X(S)= 412+/-77 dias e o dos machos de X(S)= 385+/-74 dias, o que demonstra haver uma geracao por ano. Na Tabela I, onde sao apresentados os resultados referentes a duracao de cada fase do desenvolvimento, observa-se que o tempo requerido foi crescente ate o 4 estadio, decaindo do 5 ate a fase de adultos, com as femeas seguidas dos machos. A observacao dos adultos foi realizada ate a constatacao da morte. As ninfas de 1 estadio realizaram o primeiro repasto entre o 1 e 17 dias, com maiores incidencias no 2 (15%), 4, 11 e 12 dias (12%). As alimentacoes subsequentes ocorreram no 7 e 8 dias (8%); 3, 5 e 9 dias (6%) e 1, 6, 10, 13 e 17 dias (3%). O numero de repastos por especime, durante todo o ciclo de vida, distribuiu-se homogeneamente entre um minimo de 10 (uma femea) e maximo de 32 (duas femeas). As femeas efetuaram, em media, maior numero de repastos, seguidas pelos machos e pelas ninfas de 4, 3, 5, 2 e 1 estadios (Tabela II). O intervalo de tempo entre o oferecimento do repasto e o inicio da alimentacao aumentou, gradativamente, de acordo com a evolucao do estadio ninfal, decaindo na fase adulta (Tabela III). A duracao do repasto (Tabela IV) foi crescente do 1 ao 4 estadios, seguidos por machos, femeas e 5 estadio. A quantidade de sangue ingerida (Tabela V) foi crescente ate o 3 estadio, seguido por machos, 4 estadio, femeas e 5 estadio; nao foi observada diferenca desse parametro entre as ninfas que evoluiram para qualquer dos dois sexos. Os resultados da Tabela VI demonstram que, na fase ninfal, a perda de peso foi sensivelmente maior no periodo de 24 h apos o repasto; ao contrario da fase adulta, quando a maior perda ocorreu no periodo entre 24 h apos um repasto e o proximo. O parametro defecacao foi observado no periodo de 10 min. apos o repasto, estimando-se ser este o tempo necessario para o triatomineo deixar a fonte alimentar. Desconsideraram-se os resultados referentes a fase adulta, porque o tempo requerido foi sensivelmente maior e nem todos os especimes puderam ser observados. Dos 509 repastos realizados do 1 ao 5 estadios, apenas duas ninfas (0,4%) defecaram sobre a fonte alimentar, 19 (3,7%) logo apos o repasto, porem fora da fonte e 146 (28,6%) no periodo estabelecido de 10 min. Os 342 (67,2%) restantes nao defecaram naquele periodo. Observou-se que nas ninfas de 1, 2, 3, 4 e 5 estadios os indices de defecacao foram, respectivamente, 10%, 9%, 7%, 4% e 2%. Na fase ninfal, no periodo de 10 min., obtiveram-se os seguintes resultados: 2 e 8 dejecoes - 3%; 9 - 6%; 7 - 9%; 3 e 6 - 18%; 4 - 21% e 5 - 24%. A media de defecacao dos 34 especimes neste periodo de desenvolvimento foi de X(S)= 5 +/- 2. As ninfas que originaram femeas e machos apresentaram resultados semelhantes: X = 4,94 +/- 1,85 e X = 5,12 +/- 1,65, respectivamente. DISCUSSAO Os diferentes aspectos do comportamento dos triatomineos tem sido estudados por diversos autores, especialmente aquelas especies que participam ativamente na transmissao da doenca de Chagas. Os resultados obtidos neste experimento diferem dos encontrados por Heitzmann-Fontenelle (1972) e Perlowagora-Szumlewicz (1976), possivelmente devido a variacao genetica entre as populacoes utilizadas, ao tempo em que a colônia foi mantida em laboratorio e as diferentes condicoes em que foram realizadas as observacoes. Heitzmann-Fontenelle (1972) estudou o ciclo biologico de T. pseudomaculata utilizando sangue do homem e do coelho (Oryctolagus cuniculus) como fonte alimentar. A alimentacao em ave e camundongo foi tentada, mas sem sucesso porque, no primeiro caso, os triatomineos nao se limitavam a area depenada e no segundo caso, a inquietude do camundongo impossibilitava o repasto. No presente trabalho a alimentacao foi realizada em ave e sem dificuldade. Quanto a duracao do ciclo de vida das formas jovens, os resultados do presente trabalho situaram-se entre aqueles obtidos por Heitzmann-Fontenelle (1972) e Perlowagora-Szumlewicz (1976); esta ultima autora utilizou o sangue de galinha (Gallus gallus) como fonte alimentar. Na fase adulta, ambas as autoras observaram que os machos vivem mais do que as femeas, discordando dos resultados aqui obtidos. Na fase ninfal, os especimes que originaram femeas e machos desenvolveram-se no periodo de X(S)= 68,41 +/- 29,71 e X(S)= 64,14 +/- 28,27 dias, respectivamente. Juarez (1970a) supoe que o rapido desenvolvimento dos machos possa ser atribuido a necessidade de fecundacao precoce das femeas, visando a manutencao da especie. A procura pelo primeiro repasto por ninfas do 1 estadio, segundo Perlowagora-Szumlewicz (1976), ocorreu apos o 7 dia, enquanto que para Heitzmann-Fontenelle (1972) a maioria ocorreu entre 15 e 20 dias, com o minimo de 9 e maximo de 26 dias. No presente trabalho estes dados diluiram-se entre o 1 e 17 dias, com maior incidencia no 2 dia (15%) seguido do 4, 11 e 12 dias (12%), respectivamente. Segundo varios autores, um unico repasto e suficiente para que ocorra a ecdise, exceto no 5 estadio quando ha necessidade de mais de um repasto (Brasileiro e Perondini 1974). Discordando destes autores, e de acordo com Heitzmann-Fontenelle (1972), os resultados aqui apresentados conferem mais de um repasto para cada estadio, o que pode ser justificado pelas diferentes temperaturas e fontes de alimentacao (Juarez 1970b) e pela quantidade de sangue ingerida, necessaria para que ocorra a ecdise (Heitzmann-Fontenelle 1976). As femeas, para suprirem a menor quantidade de sangue ingerida, procuraram a fonte alimentar em um curto espaco de tempo e realizaram um maior numero de repastos. Do ponto de vista epidemiologico, o tempo do repasto e importante na medida em que possibilita um contato maior entre vetor e hospedeiro, proporcionando condicoes para que a defecacao ocorra ainda sobre o mesmo, assim como aumentando as chances de o triatomineo adquirir a infeccao, especialmente ao se alimentar em um hospedeiro chagasico crônico. Segundo Heitzmann-Fontenelle (1976) esse parametro variou no periodo ninfal entre 15 e 30 min., tempo este maior do que o obtido no presente trabalho. De acordo com a mencionada autora, o tempo requerido pelos adultos foi maior porque para o repasto completo sugavam pequenas quantidades em varias areas, raramente ficando repletos, ao contrario do observado neste trabalho, cujo tempo foi menor do que o da ninfa de 5 estadio. Krinsky (1985), estudando o tempo do repasto de outras especies, verificou que as femeas realizam repasto mais longo, porem menor do que o das ninfas de 5 estadio, conforme os resultados aqui apresentados. Os resultados referentes a quantidade de sangue ingerida foram sensivelmente menores do que os obtidos por Perlowagora-Szumlewicz (1976), possivelmente devido a idade da colônia e/ou a manipulacao excessiva dos especimes (Tolezano et al. 1984b, Costa e Jurberg 1990). Comparando com os resultados obtidos com outras especies (Zeledon et al. 1970), T. pseudomaculata ingere uma menor quantidade de sangue, demonstrando nao ser uma especie expoliativa, o que nao interfere na capacidade de adquirir a infeccao (Perlowagora-Szumlewicz e Muller 1982). Nao foi observada diferenca na quantidade de sangue ingerida entre as ninfas que evoluiram para adulto, machos e femeas, o que discorda dos resultados obtidos por Perlowagora-Szumlewicz (1976) e Tolezano et al. (1984a) em relacao a T. infestans. Na fase ninfal, a perda de peso foi sensivel no periodo de 24 h apos o repasto, concordando com as observacoes de Lent e Valderrama (l977) e Cirano e Zeledon (1964) para outras especies. A frequencia com que ocorre a defecacao, durante e apos a alimentacao, e de interesse epidemiologico e demonstra a capacidade vetorial da especie que, de acordo com a literatura (Piesman e Sherlock 1983, Zarate et al. l984, Goncalves et al. 1988), pode variar entre as especies. Heitzmann-Fontenelle (1972) verificou que T. pseudomaculata so defeca sobre o hospedeiro quando atinge a replecao; quando o faz, gira o corpo 80, eliminando as fezes proximas ao local da picada e aumentando a chance de transmissao do T. cruzi. No presente trabalho este comportamento nao foi observado e o indice de defecacao decresceu nas fases ninfais, demonstrando que, quanto mais jovem for a fase, maior o numero de defecacoes; consequentemente, sao parcas as chances de adquirir e transmitir a infeccao (Torres 1915). Para T. pseudomaculata, o parametro defecacao nao se mostrou relevante em relacao a sua competencia vetorial, porque a perda de peso das ninfas ocorreu na maior parte das vezes apos os 10 min. de observacao, porem no periodo de 24 h seguintes ao repasto, e entre os adultos apos o periodo de 24 h seguintes ao repasto, confirmando que sao remotas as chances de o hospedeiro ficar exposto as fezes infectadas. De acordo com Piesman e Sherlock (1983) existem outros fatores importantes que contribuem para o estabelecimento da capacidade vetorial: a densidade domiciliar, a afinidade com o hospedeiro e o grau de adaptacao ao domicilio humano. A alta temperatura acelera o processo de alimentacao (Wood 1951) e de defecacao (Maddrell 1964), fatores de extrema importancia do ponto de vista epidemiologico. Tendo em vista que T. pseudomaculata tem sua ocorrencia assinalada em areas de clima quente, possivelmente, na natureza, o seu comportamento frente a esses parametros apresente algumas diferencas. Os estudos sobre a biologia dos triatomineos possibilitam conhecer parametros que podem contribuir para os trabalhos de controle e manutencao de colônias em laboratorio com os mais variados fins. Embora seja possivel reproduzir as condicoes do habitat natural no laboratorio, devem ser procuradas metodologias mais adequadas que interfiram menos nos resultados. Perlowagora-Szumlewicz (1976) demonstra a influencia da geracao utilizada e Tolezano et al. (1984 b) e Costa e Jurberg (1990) os efeitos do estresse causado pela manipulacao dos insetos. REFERNCIAS de triatomineos domiciliados na regiao nordeste do Brasil. Rev Saude Pub S Paulo 15: 113-164. times of insect vectors in transmission of Chagas' disease. J Econ Entomol 44: 52-54. Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Line drawing images[oc97054e.gif] [oc97054c.gif] [oc97054a.gif] [oc97054f.gif] [oc97054b.gif] [oc97054d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}