 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Diffusion model applied to postfeeding larval dispersal in blowflies (Diptera: Calliphoridae) RC Bassanezi/+, MBF Leite*, WAC Godoy**, CJ Von Zuben***, FJ Von Zuben ****, SF dos Reis *****/+/++
Departamento de Matematica Aplicada, IMECC, Universidade Estadual de
Campinas, SP, Brasil +Research Fellow CNPq This work was supported by grants from FAPESP (No 94/3851-9, 94/5355-9, 95/8031-2, 95/9299-9). Received 14 June 1996
Code Number:OC97055
Sizes of Files:
Text: 25.7K
Graphics: Line drawings and formulae (gif) - 295.6K (24 files)
This paper presents a diffusion model of larval dispersal especifically designed to account for particular aspects of postfeeding larval dispersal from the food source in organisms such as blowflies. In these organisms the dispersal of immatures includes two groups of individuals, those that are actively migrating and those that have initiated the pupation process. The classical diffusion equation in one dimension was modified to incorporate a function which describes the burying of larvae to become pupae. The analytical solution of this equation predicts oscillatory and monotonic dispersal behaviors, which are observed in experimental populations of blowfly species. Key words: Chrysomya megacephala - Chrysomya putoria - Cochliomyia macellaria - diffusion model - larval dispersal - blowflies Diffusion models have been extensively employed to investigate dispersal and have yielded considerable insight into the dynamics of animal movement in space and time (Okubo 1980, Kareiva 1982, 1983, Andow et al. 1993, Christie et al. 1995). Diffusion models can be written in the simplest form as
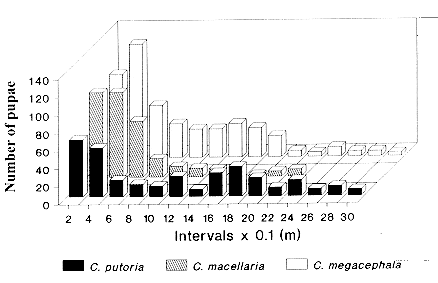
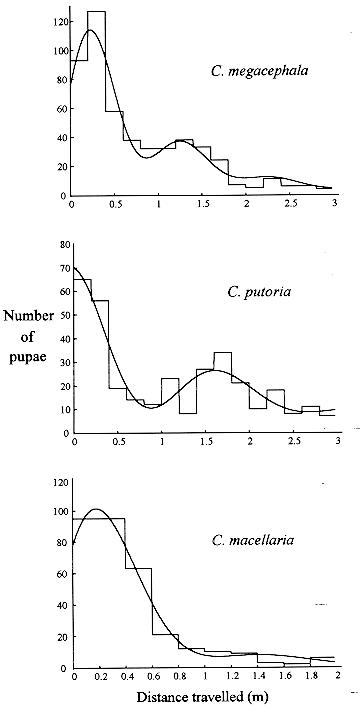
where the operator (reverse triangle) denotes the spatial gradient, t is time, u(x,y,t) is the local population density in the spatial variables x and y, D is the coefficient of diffusion, and f(u) is the reaction term describing the net population change due to birth and death. Whereas equation (1) entails several simplifying assumptions about the dispersal process (Andow et al. 1993), the ease with which distinct behavioral details can be incorporated has turned diffusion models into a powerful tool (Holmes et al. 1994). Equation (1) has been variously modified to take into account animal movement toward external stimuli or movement driven by wind or water currents (Helland et al. 1984, Banks et al. 1988), correlation of movement between directions of travel (Holmes 1993), and cases where organisms have a tendency to move away from conspecifics (Gurney & Nisbet 1975, see review in Holmes et al. 1994). Most studies employing diffusion equations have described movement in adult animals (Andow et al. 1993), but this approach has seldom been used to study dispersal in immatures, whose dispersal has important consequences for the population and community dynamics and genetic structure of populations (de Jong 1979, Roughgarden et al. 1988, Gaines & Bertness 1993). However, the same theoretical results concerning diffusion in adults can be applied to study dispersal in immatures (Broadbent & Kendall 1953). In the present study a diffusion approach is employed to model the form of larval dispersal from the food source based on experimental data from three blowfly species, Chrysomya megacephala, C. putoria, and Cochliomyia macellaria. We take advantage of the flexibility of diffusion models to incorporate behavioral characteristics of dispersal in the larval stage. The model includes the classic diffusion and velocity terms and also a function which describes the process of larvae burying in the substrate to become pupae. The focus on these blowflies is motivated by the fact that the two Chrysomya species, which are native from the Old World and Australasia, are part of an ongoing invasion of the Americas (Guimaraes et al. 1978, Baumgartner & Greenberg 1984). The invading flies have dispersed rapidly throughout the continent and have apparently caused the decline in population numbers of the native species, C. macellaria (Guimaraes et al. 1979, Guimaraes 1984, Greenberg & Szyska 1984). MATERIALS AND METHODS The process of larval migration from the food source was investigated in laboratory populations of C. megacephala, C. putoria and C. macellaria. Laboratory colonies were founded from specimens collected in the vicinities of the Universidade Estadual de Campinas, SP, Brasil, and these individuals were considered the parental generation. Adult flies were maintained at 25+/-1 C in cages (30x30x48 cm) covered with nylon and were fed water and sugar ad libitum. Eggs were obtained by providing females with fresh beef liver. Newly hatched larvae, the F1 generation, were transferred to vials (7.2 cm diameter, 13.8 cm height), containing 100 g of rearing media (Leal et al. 1982). Horizontal dispersal of full grown larvae was separately monitored for each species, depositing the vials at one end of a cardboard box (3.0 cm long, 12 cm height, 30 cm wide), covered with wood shavings, 5cm depth at the bottom. Dispersal was quantified (in cm) as the distance travelled by the larvae from the starting point at one end of the box to the point where they buried to pupate. Two replicates were run, with the following numbers of larvae in each replicate: C. megacephala, 220, 294; C. putoria, 119, 214; C. macellaria, 200, 124. RESULTS Larval dispersal - The process of larval dispersal of postfeeding larvae from the food source in calliphorids such as C. megacephala, C. putoria, and C. macellaria involves individuals that are actually dispersing while some of them begin to bury in the substrate to pupate. Because of this characteristic the form of dispersal can only be inferred when all individuals have pupated. For each species, after all individuals had pupated in the substrate, we counted the number of pupae observed at each 20 cm interval from the release point at one end of the cardboard box. Individual dispersal distances were then measured as the distance travelled by the postfeeding larvae from the point of release to the site of pupation. The number of pupae found at each interval was plotted against distance travelled by the larvae and the overall pattern that emerges from this data set is similar for the three species in that they did not move far from the food source, and the number of larvae dispersing decreases with distance (Fig. 1). Nevertheless the two invading species differ remarkably from the native species in the shape of dispersal. In C. megacephala and C. putoria, the shape of dispersal shows a pattern of dampened oscillations, whereas in the native species, C. macellaria, the number of dispersing larvae apparently declines monotonically (Fig. 1). Figure 1: horizontal dispersal in Chrysomya putoria, Cochliomyia macellaria and Chrysomya megacephala.
Diffusion model - The diffusion model developed here takes into account both dispersing individuals and those that have already buried, and is written in one-dimensional space and time as:
where c(x,t) is the concentration of larvae on surface, x is the one-dimensional spatial coordinate, t is time, D is the diffusion coefficient representing the random movement of the larvae and a is a coefficient of proportionality describing the velocity which is taken to be characteristic of each species. The first two terms in equation (2) describe the movement of larvae in the surface of the substrate. The last term in equation (2) is of particular importance in the context of larval migration because it is a function describing the rate of burying of the larvae. As an approximation to the shape of spread of larvae shown in Fig. 1, the process of larval burying in the substrate is considered to decline exponentially with the distance dispersed by the larvae and to be directly proportional to the difference in concentration of the larvae. These assumptions then lead to
where k is a constant of proportionality, c0 is proportional to the initial concentration of larvae, x* is the maximum distance travelled by the larvae, and t* indexes the end of dispersal when no larvae remain in the surface. Notice that c(x,t)>0 and h(x,t)>0 for t>t*. On the other hand, t>t*, c(x,t) = h(x,t) = 0. The concentration of buried larvae at any point in time, c*(x,t), is given by
Using equation (3) in equation (4) we obtain
When t > t*, there are no larvae left in the substrate. In this case,
describes the end of the process of larval dispersal as a result of all larvae having buried in the substrate, which implies From equations (2) and (7), Since the derivatives in equation (8) do not depend on time we have The solution of the homogeneous part of equation (9) is of the form and the roots (li) are given by the characteristic equation where p = a/D and q = k/D. The real parts of lambda 1 and lambda 2 are always negative, yielding solutions with exponential decay. The case p2 < 4q is relevant for the understanding of the pattern of dispersal because the associated complex roots lambda 1 and lambda 2 describe the damped oscillations observed in our experiments of larval dispersal (Fig. 1). The discriminant in the characteristic equation establishes a relationship between the coefficients of diffusion, D, the velocity of dispersal, a, and the constant of proportionality k, as follows. If p2 < 4q we have that
For the case where inequality (10) holds, the homogeneous solution to equation (9) can be written as
A particular solution to (9) can be found as
Substituting cp(x) in equation (9) we obtain
and
The general solution to equation (8) is thus found as
Finally, using equation (13) in equation (6) we arrive at an expression for the distribution of larvae at equilibrium, that is, when all larvae have buried to pupariate and dispersal in the substrate has ended, DISCUSSION Insects whose immature stages migrate from the food source in search of pupation sites, characterize a particular system with two populations; one of actively migrating larvae and the other of larvae burying to pupate. The process of larval dispersal is completed when all larvae have buried and no larvae remain dispersing in the substrate. These fundamental aspects were taken into account in our model by adding to the diffusion equation a function, h(x,t), describing the process of larval burying in the substrate. This function involves biologically important parameters such as the initial concentration of larvae, the maximum distance travelled by the larvae during dispersal and the time taken for dispersal to occur. The form of this function was inferred from the experimental data presented here that suggests that the decrease of the number of pupae with distance from the source is exponential.
The diffusion equation incorporating the burying function yielded an analytical solution (equation 14), which describes the equilibrium distribution of larvae buried in the substrate. This analytical solution was used to compare the dispersal behavior predicted by the diffusion model with the observed data for C. megacephala, C. putoria, and C. macellaria. Equation 14 was fitted to the data using a nonlinear regression whose details are described in the Appendix. Fig. 2 shows the fit of equation 14 to the dispersal data for the three species and demonstrates that the equilibrium distribution given by the diffusion model is consistent with the observed pattern of larval dispersal in the blowflies analyzed here. In other words, the diffusion equation incorporating the burying function seems to account for both the oscillatory and monotonic decrease in the number of pupae with increasing distance from the food source. The complexity in behavior of dispersal as indicated by the inequality in (10) is apparently the outcome of an interaction between the velocity (alpha 2), the random movement of larvae in these blowflies indexed by the diffusion coefficient (D), and also involves the constant of proportionality (k) governing the rate at which larvae bury in the substrate. Diffusion models have been extensively used to address ecological problems (Okubo 1980, Murray 1989, Holmes et al. 1994) and have been particularly useful to analyze dispersal in adult insects (Kareiva 1983, Andow et al. 1993). Nevertheless this approach apparently has not been explored to understand the dispersal behavior of immature insects such as blowflies, where two groups of individuals are interacting, those that are actively migrating and those that have already buried in the substrate. The present effort is a first approximation to the complex problem of larval dispersal, which includes factors acting at the individual level such as the minimum weight necessary for pupation (Ullyett 1950) and those intervening at the population and community levels including the timing and the velocity with which larvae leave the substrate (Peschke et al. 1987, Blackith & Blackith 1990), the spatio-temporal patterning of predation and parasitism (Norris 1959, Putman 1977, Peschke et al. 1987, Sereno & Neves 1993), the physical makeup of the substrate, and the availability of suitable places to pupate (Kneidel 1984). The application of diffusion theory has a rich tradition of illuminating mechanisms and processes in population biology (Kareiva 1983), and we believe that the approach presented here follows this tradition. As in other studies (e.g. Kareiva 1983, Possingham & Roughgarden 1990), simplifying assumptions were made such as the invariance of diffusion coefficients with space, time, and larval density. Nevertheless, the primary purpose of the model presented here was to develop a framework to analyze dispersal of immature insects living in carrion or other ephemeral substrates. The simple passive diffusion model approach taken here does include two relevant features in that it explicitly accounts for the dispersing life-stage and assumes that the dispersal distances are limited, which is commonly regarded as a limitation in diffusion models such as those applied to the dispersal of adult insects (Andow et al. 1993). We believe that the model presented here may prove a useful tool to explore the diffusion dispersal dynamics of organisms such as blowflies and other diptera where the same phase of the dispersing cycle in immatures has two stages, i.e., one actively moving and the other burying in the substrate. ACKNOWLEDGMENTS To two anonymous reviewers for comments that helped to improve the clarity of the manuscript. REFERENCES 255. APPENDIX Nonlinear Parametric Regression to Estimate the Parameters of Equation (14) The parameters presented in equation (14) can be grouped and renamed to produce the following nonlinear regression model:
For the three species C. megacephala, C. putoria and C. macellaria, the objective is to determine the corresponding set of parameters {a1, ..., a7} from the experimental data (xi,yi), i=1, ..., N. Here, x (the explanatory variable) denotes the distance travelled, y (the response variable) denotes the corresponding number of buried larvae, and N is the number of discrete distances considered (Fig. 1). The set of parameters {a1, ..., a7} can be estimated through nonlinear parametric regression, generally described as the minimization of the mean square error:
We are now prepared to estimate the set of parameters {a1, ..., a7} that minimizes J() by means of the following iterative algorithm: Initialization: Set random initial values to {a1, ..., a7} and an arbitrary small value to epsilon;
Given the experimental data, the application of this algorithm for each species gives: C. megacephala: a1 = 60.0872; a2 = 5.9472; a3 = 1.8544; a4 = 1.4474; a5 = 93.2371; a6 = 26.6265; a7 = 1.1573 C. putoria: a1 = 34.8800; a2 = 3.6839; a3 = 0.1634; a4 = 0.9026; a5 = 35.7570; a6 = 13.9524; a7 = 0.6872 C. macellaria: a1 = 85.6961; a2 = 4.6069; a3 = 1.6826; a4 = 2.3604; a5 = 87.6527; a6 = 17.6138; a7 = 1.9009 Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Line drawing images[oc97055o.gif] [oc97055m.gif] [oc97055q.gif] [oc97055j.gif] [oc97055g.gif] [oc97055c.gif] [oc97055e.gif] [oc97055b.gif] [oc97055s.gif] [oc97055p.gif] [oc97055l.gif] [oc97055d.gif] [oc97055w.gif] [oc97055k.gif] [oc97055t.gif] [oc97055a.gif] [oc97055i.gif] [oc97055f.gif] [oc97055n.gif] [oc97055h.gif] [oc97055v.gif] [oc97055r.gif] [oc97055u.gif] [oc97055x.gif] |
| |||||||||

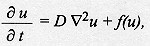{kind=link}
{kind=link}
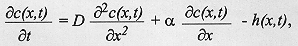{kind=link}
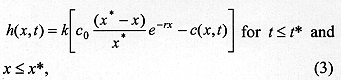{kind=link}
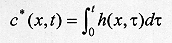{kind=link}
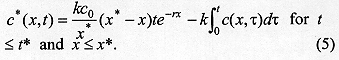{kind=link}
{kind=link}
{kind=link}
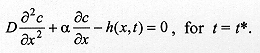{kind=link}
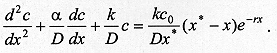{kind=link}
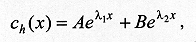{kind=link}
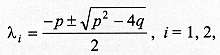{kind=link}
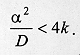{kind=link}
{kind=link}
{kind=link}
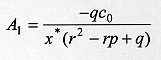{kind=link}
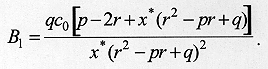{kind=link}
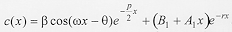{kind=link}
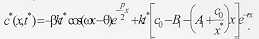{kind=link}
{kind=link}
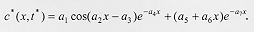{kind=link}
{kind=link}
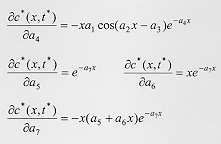{kind=link}
{kind=link}