 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
A simple reverse transcription-polymerase chain reaction for dengue type 2 virus identification Luiz Tadeu M Figueiredo/+, Weber Chelli Batista, Akira Igarashi* Unidade Multidisciplinar de Pesquisa em Virologia, Faculdade de Medicina
de Ribeirao Preto, Universidade de Sao Paulo, 14049-900 Ribeirao Preto, SP,
Brasil
+Corresponding author. Fax: +55-16-633.6695 Received 28 November 1996; Accepted 3 February 1997
Code Number:OC97072
Sizes of Files:
Text: 17.8K
Graphics: Tables (gif) - 34.7
Line drawings and photographs (jpg) - 24.1
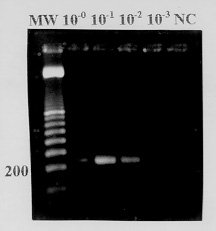
We show here a simplified reverse transcription-polymerase chain reaction (RT-PCR) for identification of dengue type 2 virus. Three dengue type 2 virus strains, isolated from Brazilian patients, and yellow fever vaccine 17DD, as a negative control, were used in this study. C6/36 cells were infected with the virus, and tissue culture fluids were collected after 7 days of infection period. The RT-PCR, a combination of RT and PCR done after a single addition of reagents in a single reaction vessel was carried out following a digestion of virus with 1% Nonidet P-40. The 50ul assay reaction mixture included 50 pmol of a dengue type 2 specific primer pair amplifying a 210 base pair sequence of the envelope protein gene, 0.1 mM of the four deoxynucleoside triphosphates, 7.5U of reverse transcriptase, and 1U of thermostable Taq DNA polymerase. The reagent mixture was incubated for 15 min at 37 C for RT followed by a variable amount of cycles of two-step PCR amplification (92 C for 60 sec, 53 C for 60 sec) with slow temperature increment. The PCR products were subjected to 1.7% agarose gel electrophoresis and visualized with UV light after gel incubation in ethidium bromide solution. DNA bands were observed after 25 and 30 cycles of PCR. Virus amount as low as 10^2.8 TCID50/ml was detected by RT-PCR. Specific DNA amplification was observed with the three dengue type 2 strains. This assay has advantages compared to other RT-PCRs: it avoids laborious extraction of virus RNA; the combination of RT and PCR reduces assay time, facilitates the performance and reduces risk of contamination; the two-step PCR cycle produces a clear DNA amplification, saves assay time and simplifies the technique. Key words: dengue identification - dengue diagnosis - reverse transcription-polymerase chain reaction The four dengue virus serotypes, designated 1, 2, 3, and 4, are positive sense, single-stranded RNA viruses belonging to the genus Flavivirus of the family Flaviviridae (WHO 1986). According to strict epidemiological criteria, dengue viruses are arboviruses (arthropod-borne viruses) transmitted by Aedes aegypti and Ae. albopictus mosquitoes (Wengler 1991). These viruses cause an important arboviral disease of man, dengue hemorrhagic fever and dengue shock syndrome (DHF/DSS) (Gubler 1987). Since the mid-80s, dengue epidemics have occurred throughout Brazil, producing millions of infections. Dengue types 1 and 2 have been isolated during epidemics. The vast majority of the disease cases involved were not life-threatening. However, after the introduction of dengue type 2, hundreds of DHF/DSS cases have occurred in the states of Rio de Janeiro and Ceara causing several deaths (Figueiredo et al. 1995). The identification of Brazilian dengue virus isolates has been done by using type-specific monoclonal antibodies in an immunofluorescence test utilizing infected mosquito cells fixed on microscope slides (Henchal et al. 1983). Reverse transcription (RT) followed by polymerase chain reaction (PCR) has been used for rapid and specific detection of flaviviruses including the dengue viruses (Shibata et al. 1990, Laille et al. 1991, Lanciotti et al. 1992, Morita et al. 1994, Beaty et al. 1995). We show here a simplified RT-PCR for detection and identification of dengue type 2 virus. MATERIALS AND METHODS Virus strains - The three dengue type 2 virus strains, CEA-2462, SPH125367 and TOC-213 (Table I), used in this study, were isolated from Brazilian patients, and kindly supplied by Dr Amelia Travassos da Rosa from Evandro Chagas Institute, Brazilian Health Ministry, Belem, State of Para, and Dr Luiza Teresinha Madia de Souza from Adolpho Lutz Institute, State of Sao Paulo Health Ministry. Yellow fever vaccine virus 17DD (Fiocruz, Brazil), was used as a negative control. C6/36 (Ae. albopictus) cells were grown at 28 C in Leibovitz L-15 medium containing 6% heat-inactivated fetal bovine serum, 10% tryptose phosphate broth, 100 U/ml penicillin and 100ug/ml streptomycin. The cells were infected with virus strains, and tissue culture fluids were collected as virus seeds after 7 days of infection with confirmation of the infection by indirect immunofluorescence (Henchal et al. 1983, Figueiredo 1990). Infected tissue culture fluids were stored at -70 C, and tenfold dilutions were prepared in tissue culture medium at time of the assay. Titration of dengue type 2 CEA 2462 - C6/36 cells were added to 96-well cluster dishes (Corning, USA) in 100ml of tissue culture medium at a density of 2x10^4 cells per well. After 24 hr the cells were infected with 50ul of tenfold dilutions of dengue type 2 CEA 2462, starting from 10^-0 to 10^-5, with 6 wells for each virus dilution. After a 5 day incubation period, all the wells received 50ul of neutral buffered formalin (pH 7) and were held overnight at 4 C. An enzyme immunoassay was performed after washing the microplate wells twice in phosphate buffered saline (PBS; Gibco BRL, USA). 100ul of dengue type 2 mouse polyclonal antiserum diluted 1/100 in PBS containing 0.5% bovine serum albumin (BSA) was added to the wells. After 1 hr at 37 C and 3 washes with PBS, horseradish peroxidase-conjugated goat anti-mouse IgG (Sigma, USA) diluted 1/2000 in 0.5% BSA-PBS was added to the wells. Microplates were maintained for 1 hr at 37 C and washed 5 times in PBS. Results were read spectrophotometrically (414 nm), 30 min after the addition of ABTS substrate (Kirkegaard and Perry, USA) to the microplate wells (Figueiredo & Shope 1987). Infected wells were determinated as positives when their OD exceeded 3 SD of the mean value for control wells in the assay. The virus titer (TCID50) was calculated by the method of Reed and Muench (1938). Primers - Dengue type 2 specific primers amplifying a 210 base pair sequence of the envelope protein gene, were synthesized by Gibco BRL (USA) (Deubel et al. 1988, Morita et al. 1993, Tanaka 1993). Primer sequences and position in the virus genome are shown in Table II. RT-PCR - The test was done in 500ul Eppendorf tubes by adding 1ul of infected tissue culture fluid (different dilutions were used), and 5ul of a detergent mixture containing 1% Nonidet P-40 and 5 U of RNase inhibitor (Pharmacia, USA) in PBS. After 60 sec of incubation at room temperature for virus solubilization, the cDNA was added to a reaction mixture containing 50 pmol of the primer pair, 0.1 mM of the four deoxynucleoside triphosphates, 7.5 U of reverse transcriptase (Pharmacia, USA), 1U of Taq DNA polymerase (Pharmacia, USA), and 5ul of a buffer solution containing 5 mM Tris (pH 9.0), 0.75 mM MgCl2, and 25 mM KCl. A total volume of 50ul was obtained by adding distilled and deionized water. The reaction mixture was covered with 2 drops of oil and was incubated for 15 min at 37 C for RT, followed by 15 to 30 cycles of a two step PCR amplification (92 C for 60 sec, 53 C for 60 sec) in a thermal cycler (Techne, UK). The temperature increment was slow, taking 120 sec from 53 C to 92 C, and 15 sec from 70 C to 75 C. The total time for the test was 150 min. Eight ul of each PCR product was subjected to electrophoresis in a 1.7% agarose gel. PCR product nucleotide was visualized in UV light after incubation of the gel in a 0.5ug/ml ethidium bromide solution for 20 min, and rinsing with tap water. The size of the amplified DNA fragments was determined by comparing their migrations in the gel with those of the bands of a 100 base pair molecular weight marker (Pharmacia, USA). RESULTS The undiluted dengue type 2 CEA 2462 seed was processed at 20, 25 and 30 cycles of PCR after RT, in order to determine the ideal number of amplifications for the test. DNA bands of 210 base pairs were observed after 25 and 30 cycles. From this result, 30 cycles was considered as suitable for the test. For evaluation of sensitivity of the RT-PCR, the 10^4,8 TCID50/ml dengue type 2 CEA 2462 virus seed was tested at serial tenfold dilutions. Amplified 210 base pair DNA bands were observed at 10^-2 (weak band), 10^-1, and 10^-0 dilutions (Fig. 1).
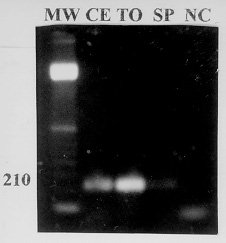
NC: negative control ( RT-PCR product of yellow fever virus). Figure 2: photograph of an agarose gel stained with ethidium bromide, showing the specific DNA amplification by reverse transcription-polymerase chain reaction of the three dengue type 2 virus seeds at 10-1 dilution. Yellow fever virus was used as a negative control. DNA amplification was not observed in some assays when undiluted virus seeds were processed, though a clear DNA amplification was obtained with the same virus seed at 10^-1 dilution (Fig. 2). Specific DNA amplification was observed with dengue type 2 CEA 2462, SPH 125367 and TOC 213 at 10^-1 dilution. DNA amplification was not observed with yellow fever virus (Fig. 2). DISCUSSION The dengue type 2 specific primers utilized in this assay have been previously studied (Tanaka 1993, Morita et al. 1993, 1994). The high specificity of the 210 base pair sequence of the envelope protein gene produced in this study was exhaustively confirmed in the papers cited (Tanaka 1993, Morita et al. 1993, 1994) eliminating the need for further confirmation by hybridization or nested PCR. Also, the absence of DNA amplification of yellow fever virus excludes the possibility of any nonspecific amplification related to the virus, the cell culture or the nutrient medium in our study. Three dengue type 2 virus seeds obtained from cells which had confirmed infection by immunofluorescence, were amplified by RT-PCR, showing a good correlation between these techniques. Two-step PCR cycles have been reported as generating as much DNA as three-step cycles, at least for fragments about 200 base pairs long or less (Cha & Thilly 1995). A suitable extension of DNA in two-step PCR is accomplished by the high processivity of taqDNA polymerase action which occurs during the temperature transition of 53 C to 92 C. The optimal Taq DNA polymerase reaction range temperature from 70 to 75 C, took 15 sec in the cycles of our RT-PCR (Cha & Thilly 1995). The 30 cycles of RT-PCR gave a very high sensitivity and a suitable amplification efficiency, since virus titers as low as 102,8 TCID50/ml could be detected. Virus titers in the blood during the first 6 days of dengue infection are commonly higher than this level, so the presence of the virus could be detected by RT-PCR (Gubler et al. 1981). However, in some tests, DNA amplification was not observed when undiluted virus seeds were processed, despite a clear DNA amplification obtained with the same virus seed at 10^-1 dilution. Water used as sample diluent in our RT-PCR probably reduces RNAse, reverse transcriptase or DNA polymerase inhibitors from the C6/36 cells, or reduces RNA excess present in straight samples inhibiting reverse transcriptase. Compared to other reported assays (Shibata et al. 1990, Laille et al. 1991, Lanciotti et al. 1992, Tanaka 1993, Morita et al. 1993, 1994, Beaty et al. 1995) our RT-PCR shows the following advantages: (i) solubilization of the virus by detergent mixture (Nonidet P-40 and RNAse inhibitor) prior to the RT-PCR, avoids laborious extraction of virus RNA especially when assaying many samples; (ii) the combination of RT and PCR which can be carried out after single addition of reagents in a single reaction vessel, reduces assay time, facilitates the performance and reduces the risk of contamination; (iii) the use of small amount of reagents (1U of Taq DNA polimerase, 7.5 U of reverse transcriptase, 50 pmol of the primer pair) indicates that the method is reasonably economical. This makes the technique applicable even in the third world countries such as Brazil; (iiii) the two-step PCR cycles produce a clear DNA amplification, saves assay time and simplify the technique. An obvious application of this RT-PCR is for confirming dengue type 2 virus isolation from patients or mosquitoes, in tissue culture fluids. We intend to extend the use of this technique to other dengue serotypes, and, since dengue outbreaks presently occur in Brazil, we will test the amplification of dengue virus genome from clinical specimens. Acknowledgements To Dr Benedito Fonseca for helpful suggestions, Mrs Rita Helena Carlucci for technical assistance, and Dr David DeJong for review of the manuscript. Research supported by Japan International Cooperation Agency, and Brazilian Agencies CNPq and CAPES. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97072c.jpg] [oc97072d.jpg]Line drawing images[oc97072b.gif] [oc97072a.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}