 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Contrasting genomic variability between clones from field isolates and laboratory populations of Schistosoma mansoni Patricia Machado Pinto/+, Cristiana FA Brito, Liana K Jannoti Passos*, Miriam Tendler**, Andrew JG Simpson*** Laboratorio de Parasitologia Celular e Molecular +Corresponding author. Fax: + 55-31-295.3115 Received 28 August 1996; Accepted 28 February 1997
Code Number:OC97075
Sizes of Files:
Text: 17.5K
Graphics: Line drawings and photographs (jpg) - 119.1K
The extent of genomic variability of clones of Schistosoma mansoni obtained from field isolates was compared with that of strains that have been laboratory maintained. Analysis was undertaken using randomly amplified polymorphic dNAs (RAPDs) generated with three primers. Phenograms showing the similarity among the clones were constructed. The data showed that while the laboratory strain is highly homogeneous the clones derived from the field populations were highly variable with 43% of RAPDs exhibiting polymorphisms among 23 clones. Clones isolated from the same infected individual were always more closely grouped than clones from different individuals. The data clearly demonstrated that earlier analyses of the genomic variability in S. mansoni have underestimated this phenomenon due to the failure to examine field isolates. Key words: polymerase chain reaction - randomly amplified
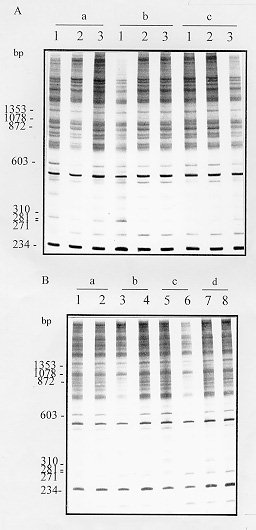
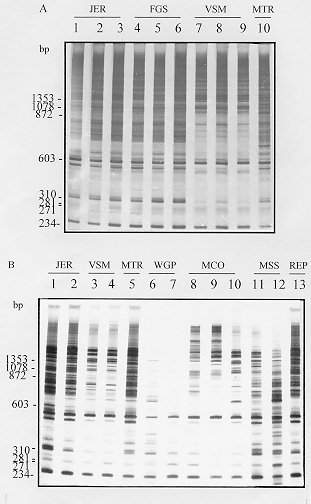
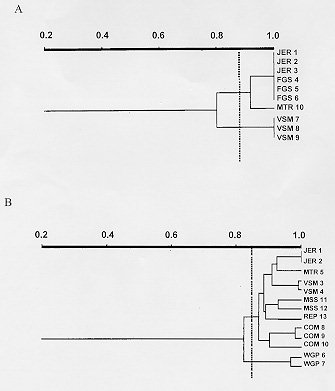
polymorphic DNA - schistosomiasis - Schistosomiasis is caused by infection with blood flukes of the genus Schistosoma, of which three species (S. mansoni, S. haematobium and S. japonicum) are the main causative agent of the disease in man (Rollinson & Southgate 1987). This chronic helminthic infection affects more than 200 million people throughout the world and 600 million live in endemic areas where they are at risk of infection (Mott 1987). This wide distribution of the disease makes the study of genomic variability extremely important. Schistosomes exhibit significant intraspecific variation as indicated by differences in snail infectivity (Paraense & Correa 1963, Richards & Shade 1987) and drug susceptibility (Katz et al. 1973, Araujo et al. 1980, Dias et al. 1982). The study of ribosomal DNA (rDNA) has shown that schistosomes exhibit both interspecific and intraspecific differences. It has been suggested that the variation in the major tandemly repeated copies of the gene can be used for species identification (Simpson et al. 1984, McCuthchan et al. 1984, Walker et al. 1986, 1989) whereas low copy numbers variants that exhibit intraspecific polymorphism can be used for strain identification (McCutchan et al. 1984, Simpson et al. 1984, Vieira et al. 1991). However, the data available from isoenzyme analysis of laboratory strains of schistosomes from Africa, Asia, the Caribbean and Brazil (Fletcher et al. 1981) indicate that there is relatively restricted diversity in these species and strains with no distinct genetic constitutions. Recent work done in our laboratory using random amplified polymorphic DNA (RAPD) showed very discrete variability in different strains of S. mansoni adult worms, however, all of them were derived from laboratory maintained populations (Dias Neto et al. 1993). To determine whether this low variability is due to the maintenance of the worms under laboratory conditions or whether it is a real characteristic of this parasite, we compared genomic variability of natural populations of S. mansoni derived from patients in Brazil with an established laboratory strain by RAPD analysis. This technique enables a large number of independent genetic loci to be examined that are not biased towards particular sequences or types of sequences and can thus be taken as representative of the genome (Williams et al. 1990, Welsh & McClelland 1991). MATERIALS AND METHODS Parasites and DNA preparation - LE strain adult schistosomes (Pellegrino et al. 1967, Araujo et al. 1980) were perfused from the mesenteric veins of 5 mice 45 days after infection. Individual worms, 8 male and 8 female, were collected from these mice, washed in physiological saline and freeze-dried at -70 C for subsequent DNA preparation. DNA was extracted from individual worms. The field isolates were obtained from the patients with schistosomiasis FGS, JER, MCO, MSS, MTR, REP, VSM and WGP from faecal samples. The patients were from an endemic area - Nossa Senhora de Fatima - Sabara, MG, Brazil. The S. mansoni eggs were isolated by faecal sedimentation, followed by filtration through gauze. The miracidia were hatched from these eggs using artificial light. Monomiracidial Biomphalaria glabrata infection was done by incubating single miracidia with single snails, in the same well of 24 wells plaques, for 6 hr at 26-28 C, producing a variable number of clones from each faecal sample. The infected snails were maintained in aquarium without light for 45 days and the cercariae recovered by exposure to artificial light. Individual adult worms and cercarial clones produced from monomiracidial infection of B. glabrata were recovered by centrifugation 2,500 rpm for 5 min. The parasites were resuspended in 50mM Tris-HCl, 50mM EDTA, 100mM NaCl, 0.5% SDS, pH 8.0 and incubated with 20 mg/ml proteinase K for 2 hr at 37 C. Following phenol extraction and ethanol precipitation the DNA was resuspended in 10mM Tris-HCl, 1mM EDTA pH 8.0 (TE). PCR amplification and polyacrylamide gel electrophoresis - Each amplification was done in a final volume of 10 ml containing 1.0 unit of Taq DNA polymerase (Cenbiot, RS, Brazil), 200 uM of each dNTP, 1.5mM MgCl2 , 50mM KCl, 10mM Tris-HCl, pH 8.5 together with 12.8 pmol of primer and either 0.1 or 1.0ng of template DNA. The primer 3302 (5'-CTGATGCTAC-3'), 3307 (5'-AGTGCTACGT-3') and lambda gt11R (5'-TTGACA CCAGACCAACTGGTAATG-3') were arbitrarily selected from laboratory stocks. The reaction mixture was overlaid with 20 ul of mineral oil and, following an initial denaturation at 95 C for 5 min, was subjected to 2 cycles through the following temperature profile: 30 sec at 95 C for denaturation, 30 C for 2 min for annealing and 72 C for 1 min for extension and followed by 33 cycles for which the annealing step was altered to 40 C. In the final cycle, the extension step was for 5 min. Following amplification, 3 ul of the reaction was mixed with DNA sample buffer (0.125% xylene cyanol, 0.125% bromophenol blue, 15% glycerol) and subjected to electrophoresis through a 4% polyacrylamide gel (acrylamide-bisacrylamide 29/1) in TBE buffer (2mM EDTA, 10 mM Tris-borate, pH 8.0). The gels were silver stained by fixing with 10% ethanol/0.5% acetic acid for 20 min, stained with 0.2% silver nitrate for 30 min, and reduced with 0.75 M NaOH/0.1M formaldehyde for 10 min (Santos et al. 1993). Data analysis - To calculate percentage band sharing, the bands observed in a given lane were compared with those in other lanes of the same gel. Enlarged photographs of the gels were examined and the principal bands of 1.5 kb or less were scored. A taxon/character matrix was constructed on the basis of the presence/absence of bands. The phenogram based on band sharing between all possible pairs in an analysis group were constructed using the Dice similarity coefficient (Dice 1945), using the formula: S = 2a/2a + b + c where a = the number of bands shared between organisms 1 and 2, b = the number of bands present in 1 but not in 2 and c = the number of bands present in 2 but not in 1. The data derived from this formula were collected into a matrix of similarity that was then used for unweighted pair group method analysis (UPGMA). The phenon line marked on the UPGMAs is the average of the similarities among the pairs and indicates the point of reference for dividing the organisms into separate groups (Sneath & Sokal 1962). RESULTS AND DISCUSSION In order to examine genomic variability in individual male and female adult worms, specimens from mice experimentally infected with S. mansoni strain LE were used to extract DNA. The DNA was amplified using primer 3302 (Figs 1A, B). The identical profiles are consistent with the low variability in laboratory strains previously reported (Dias Neto et al. 1993). Figure 1: A - Randomly amplified polymorphic DNA profiles of individual Schistosoma mansoni LE strain, males (lanes a), females (lanes b) and pairs (lanes c) from three different mice, using 1.0ng of template and primer 3302. B - Analysis of 8 individual S. mansoni, males (lanes 1, 3, 5 and 7) and females (lanes 2, 4, 6 and 8) from 4 different mice (a - d), using 1.0ng of template and the same primer (The polymerase chain reaction products were resolved by electrophoresis on a 4% polyacrylamide gel followed by silver staining). To investigate whether there is higher variability in field populations of S. mansoni adult worms, clones of cercariae were obtained from snails infected with an individual miracidium hatched in vitro from S. mansoni eggs obtained from infected patients from an endemic area. Figs 2A and B shows the RAPD profiles of cercarial clones from distinct patients, amplified with primer 3302. RAPD profiles of clones from the same patient were similar, but not identical, whereas the profiles of clones from different patients showed higher levels of polymorphisms. Similar results to these shown for primer 3302 were also obtained with primers 3307 and lambda gt11R. 43% of the bands that were amplified by arbitrarily selected chosen primers (3302, 3307 and lambda gt11R) were found to be polymorphic. Figure 2: Figure 3: A and B - Unweighted pair group method analysis phenograms of the clones obtained from infected individuals based on pairwise band sharing. Corresponding to the gels shown in Figs 2 A and B, respectively. The similarity coefficient shown on the horizontal scale was derived from the Index of Dice. The vertical bar represents the phenon line, which shows the average level of similarity between all of the pair samples analyzed. Phenograms, were composed using all of the data derived from all primers in which the most prominent and well resolved bands of 1.5kb or less were scored. A complete similarity among clones from the laboratory strain, LE, was observed with no detectable polymorphic bands. The phenograms in Fig. 3 show the analysis of RAPDs derived from the cercariae clones of the field population. The total number of bands scored was 63, with the average number of bands scored per lane being 21. The phenogram demonstrated that different clones from same individual could be grouped together. The phenon line determined the formation of two groups: one where clones from JER, FGS and MTR are grouped and the other comprising the VSM clones (Fig. 3A). The increase of samples number confirmed these results (Fig. 3B). This result provides a direct demonstration of the variation in a natural population of schistosomes from infected individuals. We observed that the genetic variation among individuals worms comprising natural populations was marked, and the variation in a natural population of schistosomes, in terms of the complexity of the RAPD profile, appeared to be significantly greater than that observed in parasites that had been maintained in the laboratory for numerous generations. The data obtained here are in agreement with those reported in studies using rRNA (Vieira et al. 1991) with natural S. mansoni populations and demonstrate that such variability is commonly found in the Schistosoma genome. It should be noted that, to our knowledge, clonal analysis of laboratory strains of S. mansoni by RAPD has not been previously undertaken although comparison of mixed populations has suggested little heterogeneity. The extreme homogeneity of LE may be due to artificial selective pressure operating during laboratory maintenance or to the fact that the strain originated from a single individual who may have had an infection of limited genetic variability. ACKNOWLEDGMENTS To Dr Alvaro Romanha from Laboratorio de Parasitologia Celular e Molecular, Centro de Pesquisas Rene Rachou-FIOCRUZ, for the facilities in the development of the project. To Dr Ana Rabello of the Laboratorio de Esquistossomose, Centro de Pesquisas Rene Rachou who provided the faecal samples from which the field isolates were derived. Supported by CNPq. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97075c.jpg] [oc97075a.jpg] [oc97075b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}