 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Incoordination, paralysis and recovery after pyrethroid treatment on nymphs III of Triatoma infestans (Hemiptera: Reduviidae) Raul A Alzogaray, Eduardo N Zerba+ Centro de Investigaciones de Plagas e Insecticidas, (CIPEIN-CITEFA/ CONICET), Zufriategui 4380, 1603 Villa Martelli, Buenos Aires, Argentina +Corresponding author. Fax: +54-1-709.3210 Received 27 May 1996; Accepted 19 February 1997
Code Number:OC97079
Sizes of Files:
Text: 18.9K
Graphics: Tables (gif) - 82.4K
Line drawings (jpg) - 34.6K
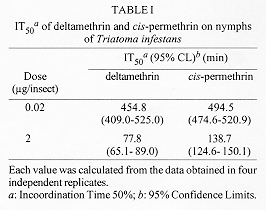
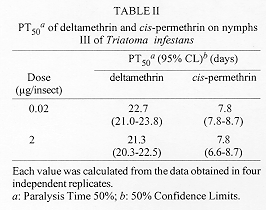
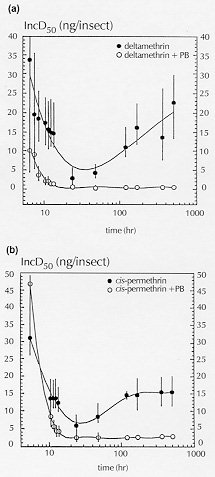
Symptoms of poisoning for deltamethrin and cis-permethrin on nymphs III of Triatoma infestans were described. The time required for incoordination and paralysis were determined. Deltamethrin was equal or more rapid in the onset of the first effect (accordingly to dose), and cis-permethrin in the onset of the second one. There were no significant differences between incoordination doses 50% (IncD50s) at different times for the two pyrethroids. They showed equivalent incoordination power, but the nymphs treated with deltamethrin recovered slightly more rapid and in greater amount than the nymphs treated with cis-permethrin. The recovery was inhibited by the simultaneus application of piperonyl butoxide. This result suggests that biotransformation by mixed-function microsomal oxidases are involved in the process of recovery. Key words: Triatoma infestans - deltamethrin - cis-permethrin - pyrethroids - poisoning symptoms The bloodsucking bug Triatoma infestans is one of the more important vectors of Chagas' disease in South America (WHO 1991), and the pyrethroids are the more effective triatomicides (Casabe et al. 1988). These neurotoxic insecticides are an effective tool in national control programs in Argentina (Zerba 1988). Particularly, deltamethrin is a cyanopyrethroid broadly used in control programs (Zerba 1989). Permethrin is an isomers mixture well established against public health pests because of its high insecticidal power and low mammalian toxicity (WHO 1990), but it is not usually used in triatomine control because of its unfavourable cost-benefit relationship. Previous works by our laboratory showed that the cis isomer of permethrin have an important insecticidal activity on nymphs I of T. infestans (Alzogaray & Zerba 1996). On the other hand, it has been demonstrated that the less active trans isomer produces an antagonistic effect which masks the high toxicity of the cis isomer when they are simultaneously applied on both nymphs III and V of T. infestans (Alzogaray et al. unpublished results). These results suggest that cis-permethrin has potential use in the control of Chagas' disease vectors. The specific aim of the present work was to compare the onset of incoordination, paralysis and recovery by deltamethrin and cis-permethrin on nymphs III of T. infestans. This knowledge could be relevant to analyze a possible use of cis-permethrin in future measures for Chagas' disease vectors control. MATERIALS AND METHODS Biological material - Nymphs III of T. infestans were obtained from a colony mantained in our laboratory at 28 C and a photoperiod of 12: 12 (L: D) hr. The experiments were done on 7-11 days old nymphs starved from the last moult. Chemicals - Deltamethrin [(S)-alpha-cyano-3-phenoxybenzyl (1R)-cis-3- (2,2- dibromovynil) -2,2- dimethylcyclopropanecarboxylate] (97%) was provided by courtesy of Roussel-Uclaf (France). Cis-permethrin [3-phenoxybenzyl (1RS)-cis-3-(2, 2-dichlorovynil)-2,2-dimethylcyclopropanecar-boxylate] (99%) was a gift from Chemotecnica Sintyal (Argentina). Symptomatology - Pyrethroids were topically applied on the ventral side of the abdominal surface in 0.2 ul of an acetonic solution, each insect receiving 0.02 ug or 2 ug of insecticide. The nature and order of appearance of the symptoms of poisoning were recorded by individual observation of at least 10 nymphs per dose. The following sequence of symptoms was observed: (1) sligth tremors; (2) backward or side displacement; (3) contraction of the three pairs of legs beneath the body; (4) elevation above the body plane; (5) paralysis of the third pair of legs; (6) violent convulsive movements; (7) paralysis of the three pairs of legs; (8) slow or occasional movements; (9) lack of movements. Bioassays - Pyrethroids were topically applied on the ventral side of the abdominal surface in 0.2 ul of an acetonic solution. Control groups were topically treated with acetone alone. Immediately after topical application the nymphs were placed in a constant temperature chamber at 26 C (60-90% RH). To calculate the time required for 50% of the experimental population to show incoordination or paralysis (IT50 or PT50, respectively), groups of 5 nymphs were treated topically with a single dose of pyrethroid in acetone. The experiments were performed with either 0.02 or 2 ug/insect. The first of these doses is approximately the incoordination dose 99% (IncD 99) for both the pyrethroids assayed. The evaluated effects were (1) incoordination, defined as the incapacity to leave a filter paper disc of 15 cm diameter in tenfold time required by controls (which is < 5 sec); and (2) paralysis, defined as lack of movements in legs, antennae and proboscis. The appearance of these effects was followed as a function of the time after the treatment. Each experiment was replicated four times. Doses between 2 and 128 ng/insect were applied to groups of five nymphs to calculate the IncD50. Nymphs were examined for incoordination at different times after insecticide application in order to establish the dose-response relationship. In this way, an IncD50 value for each time was obtained. The bioassays were interrupted at 21st day, when the control mortality was greater than 10%. Each experiment was replicated twice (N = 40-60). The values of IT50, PT50 or IncD50 were calculated using the probit method (Litchfield & Wilcoxon 1949). Differences between values were considered significant (P < 0.05) if the respective 95% CL did not overlap (Toth & Sparks 1988). Filter papers Whatman # 1 were impregnated with a solution of piperonyl butoxide in acetone (0.07 mg/cm^2). The nymphs were permanently exposed to the treated papers during each experiment. Control groups were exposed to filter papers treated with acetone alone. Incoordination and recovery analysis - IncD50 versus time curves were made to show the distinct phases described by Sawicki (1962) on the housefly Musca domestica: a falling line (phase 1), a rising line (phase 2), and a plateau (phase 3). This way to present the results allow to see how the proportion of affected insects at each dose increases as a function of the time after the insecticide application, and it reaches a minimum value (the Incoordination Endpoint - IE). After IE is reached, the curve increases while the insects recovery from the temporary incoordination. Finally, the recovery stops and a plateau is observed. The Effectivity: Incoordination Ratio and the Incoordination Recovery Rate (IRR) were calculated based in Sawicki (1962). Effectivity: Incoordination Ratio = IncD50 p3 / IncD50 ep; where IncD50 p3 is the IncD50 in the phase 3 of the curve, and IncD50 ep is the minimum IncD50 observed before recovery occurs (IE). IRR is the slope of the phase 2 of the curve and it is expressed as ng/nymph/day. The Effectivity: Inccordination Ratio is a measure of how much recovery occurs. The IRR is a measure of how rapidly the recovery occurs. RESULTS AND DISCUSSION The symptoms of intoxication by pyrethroids observed in T. infestans (see Materials and Methods) have been reported by other authors in the cockroach Periplaneta americana (Gammon et al. 1981, Benoit et al. 1985), and Musca domestica (Adams & Miller 1980). A correlation between visible and neurophysiological events of poisoning by allethrin have been established in P. americana (Gammon 1978). Results in Table I show that the values of IT50 were inversely proportional to the dose for the two pyrethroids assayed. When a dose of 0.02 ug/insect was applied, the IT50 for both pyrethroids were not significantly different (based on overlap of 95% CL, P > 0.05). In change, when the dose was 2 ug/insect, the IT50 for deltamethrin was significantly smaller than the IT50 for cis-permethrin (based on nonoverlap of 95% CL, P < 0.05). Results in Table II show the values of PT50 for the two insecticides. For the same dose, the nymphs treated with cis-permethrin showed lack of movements in a time significantly smaller than the nymphs treated with deltamethrin (based on nonoverlap 95% CL, P < 0.05). Not significant differences were observed for PT50 produced by different doses of a same insecticide (based on overlap of 95% CL, P > 0.05). Following the suggestion of Burt and Goodchild (1974) that the central nervous system is the site where the pyrethroids act to induce incoordination in insects, Briggs et al. (1976) concluded that rapid incoordination must involve fast and efficient penetration to the central nervous system. This was contrasted with kill, which can occur two days or more after application. These authors suggested that an important property determining whether a particular pyrethroid will be effective for incoordination or kill will be the overall polarity of the molecule, which they considered to have a marked influence on the rate at which the pyrethroid would penetrate into the site of action. Briggs and coworkers observed that among pyrethroids the optimum polarity for incoordination is slightly on the lipophobic side of that for kill, and suggested that rapid penetration, which they assumed the more polar pyrethroids can achieve, is required for incoordination. Clements and May (1977) suggested that knockdown by pyrethroids results from a particular type of physiological action, and that the ability to induce that action is dependent upon molecular structure. These authors demonstrated that the a-cyano moiety enhances knockdown activity of the pyrethroids. The two hypothesis above mentionated are not mutually exclusive. In fact, our results are according to both of them. Figure : Incoordination doses 50% (IncD50) versus time curves for topically applied (a) deltamethrin and (b) cis-permethrin, with or without piperonyl butoxide, on nymphs III of Triatoma infestans. The nymphs were permanently exposed to filter paper treated with a solution of piperonyl butoxide in acetone (0.07mg/cm^2) or acetone only (controls). Each value was calculated from the data obtained in two inde[endent replicates (N=40-600. Bars represent the 95% confidence limit.Figure shows the variation of IncD50 for deltamethrin and cis-permethrin as a function of the time after the insecticide application, with and without piperonyl butoxide. In the curve for cis-permethrin (Fig. b) the three phases described for pyrethrum extract in M. domestica (Sawicki 1962) were observed: rapidly falling line (phase 1), gradually rising line (phase 2), and attaining a plateau (phase 3). In the curve for deltamethrin (Fig. a), only the two first phases were observed. Before the phase 3 (a plateau) was reached, the bioassays were interrupted because of the mortality of controls was greater than 10% (at the 21st day). The comparison of IncD50 for the two insecticides in each time indicates that there were no significant differences between the effects of deltamethrin and cis-permethrin in any time (based on overlap 95% CL, P > 0.05). Table III shows the toxicological parameters calculated from the curves in Fig. For the two insecticides assayed, the IncD50 (24 hr) coincided with the IE . The IRR for deltamethrin was greater than for cis-permethrin (0.67 and 0.55 ng/nymph/day, respectively). The Effectivity: Incoordination Ratio was at least threefold for deltamethrin than for cis-permethrin (³ 8.1 and 2.7, respectively). These results mean that the two pyrethroids showed similar inccordination power, but the nymphs treated with deltamethrin recovery slightly more rapid and in greater amount than those treated with cis-permethrin. When piperonyl butoxide was applied, recovery was not observed in nymphs treated with either deltamethrin or cis-permethrin (Fig.). These results indicate that the activity of mixed-function microsomal oxidases are involved in the process of recovery. The knockdown effect is an important part of the pyrethroid action. It is rapid in onset, sometimes very long lasting, and not necessarily associated to lethal action (Naumann 1990). The three phases in the curve KD50 vs log time were described for pyrethrum and a number of pyrethroids in M. domestica (Sawicki 1962, Scott & Georghiu 1984). In these cases, the knockdown ocurred few minutes after the application of the insecticides, and the KD50 reached a plateau near the 24th hr. Our results indicate that the events of incoordination and recovery present a very different kinetic in T. infestans. Penetration, metabolic degradation, and interaction of the insecticides with the site of action are factors known to influence the knockdown (Ruigt 1985). However, it is not clear if knockdown and kill are or not different manifestations of the same underlying mechanism. The degradation at the site of action was proposed to explain the rapid recovery in M. domestica (Sawicki 1962). This hypothesis was supported by the lack of housefly recovery in the presence of piperonyl butoxide, an inhibitor of the activity of mixed-function microsomal oxidases. Similar results were reported in adults of T. infestans treated with deltamethrin and piperonyl butoxide (Casabe et al. 1988). Previous result by our laboratory (Alzogaray & Zerba 1996, Alzogaray et al., unpublished results) and the evidence presented in this paper indicate that cis-permethrin can be considered a potential tool for the Chagas' disease vector control, and more investigations in relation with its properties should be done. This investigation received financial support from the UNDP/World Mundial Bank/WHO Special Programme for Research and Trainning in Tropical Diseases, from the Consejo Nacional de Investigaciones Cientificas y Tecnicas of Argentina (CONICET), and from Chemotecnica Sintyal (Argentina). REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97079c.jpg]Line drawing images[oc97079b.gif] [oc97079a.gif] [oc97079d.gif] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}