 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Characterization of a Trypanosoma rangeli strain of Colombian origin C Zuniga, T Palau^*, P Penin^**, C Gamallo^***, JA de Diego^**/+ Departamento de Medicina Experimental, Facultad de Medicina, Campus
Occidente, Universidad de Chile, Santiago, Chile Received 9 September 1996
Code Number: OC97094
File Sizes:
Text: 22.2K
Graphics: Tables (jpg) - 50.3K
Line drawings and photographs (jpg) - 163.7K
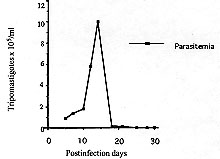
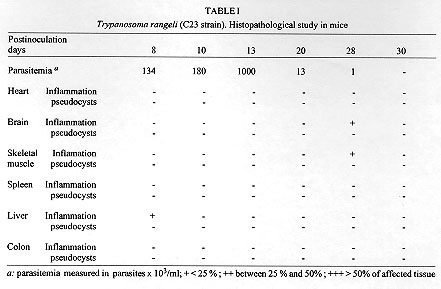
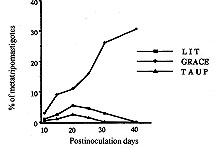
A Colombian strain of Trypanosoma rangeli was characterized by analyzing its behaviour in different axenic and cellular culture, its infection rate and the histopathological lesions produced in experimental animals. Although slight inflammatory infiltrations were shown in different histopathological sections, no pseudocysts could be observed. Grace's insect medium is better than liver infusion tryptose or artificial triatomine urine supplemented with proline when studying T. rangeli metacyclogenesis, with a peak of 32% trypomastigotes. High infection rates were found in VERO and J774 cells. Because of its 100% infectivity rates and adequacy of parasitemia levels, C23 strain is a suitable model of T. rangeli biology study. Key words: Trypanosoma rangeli - histopathological study - culture - infection rate - Colombia Trypanosoma rangeli is a trypanosome species that infects man in Central and South America where it can be found in mixed infections together with T. cruzi in both invertebrate and vertebrate hosts. While T. cruzi causes Chagas' disease, T. rangeli is considered nonpathogenic to man (D'Alessandro 1976, De Souza 1984). Mixed infections represent a serious problem to the differential diagnosis as it shares common antigenic determinants with T. cruzi (Guhl & Marinkelle 1982). Both parasites have a similar geographical distribution, the same vertebrate hosts, and, in some regions, identical insect vectors (D'Alessandro 1976, Guhl et al. 1985). Study of parasitism of this flagellate in mammalian hosts has been hampered by the difficulty of obtaining bloodstream forms in the laboratory animals with very low parasitemias and short duration in experimental animals (Urdaneta-Morales & Tejero 1986). Another important fact is the scarcity of studies about the influence of several factors in the infection such as parasite strain, inoculum dose, hosts age and sex, and immune response related to the host. This kind of studies as well as those related to the mixed infections with T. rangeli and T. cruzi in experimental animals requires T. rangeli strains with high infectious capacity able to produce high and prolonged parasitemia levels which permits its evolution study during the infection. The present paper describes the characterization of a Colombian strain of T. rangeli in several aspects such are: biological behaviour in acellular media, in vitro infections of cell line cultures and analysis of the mice infection with reference to the parasitemia evolution and the histopathological study in different tissues. MATERIALS AND METHODS Trypanosomes - Strain C23 of T. rangeli was used. It was isolated from a primate Aotus trivirgatus in 1982 in San Marcos, Sucre, Colombia and identified by polymerase chain reaction (PCR) (Campbell et al. 1993). Experimental animals - Groups of 10 male mice, one month aged, of the strain Swiss ICO NMRI (IOPS) were used to study the parasitemia evolution and the histopathological analysis. Parasitemia study - A group of 10 mice was inoculated intraperitoneally with 2.5 x 10^5 bloodstream trypomastigotes of T. rangeli from mice whose infection was previously stabilized. Parasitemias were determined daily until they were negative. For this study, blood of each mice were collected in a microhaematocrit tube (60 ul) by the method described by Arias and Ferro (1988). Histopathological study - A group of 10 mice infected with 2.5 x 10^5 parasites was sacrified at days 8, 10, 13, 20, 28 and 30 postinfection (p.i.). For each interval a study of heart, brain, skeletal muscle, liver, spleen and colon was made. Sections were fixed in formalin and embedded in paraffin and 8 um thick sections were stained with haematoxylin and eosyin. The severity of inflammatory infiltration or pseudocysts presence was graded on a 3-point scale: + indicates less of 25% of affected area; ++ between 25 and 50% of affected area; and +++: more than 50% of affected area. Study of the metacyclogenesis - Transformation studies in three acellular media: liver infusion tryptose (LIT) (Camargo 1964), Grace (Grace 1962, Sullivan 1982) and artificial triatomine urine supplemented with proline (TAUP) (Contreras et al. 1988) were made. Parasites were preincubated for 2 hr in artificial triatomine urine (TAU) (190 mM NaCl, 17 mM KCl, 2 mM MgCl2, 2 mM CaCl2, 0.035% NaHCO3, 8 mM phosphate buffer pH 6.0) prior the study in TAUP medium. An initial population of 10^4 parasites was incubated at 26^oC in an acellular culture of a total volume of 10 ml into sterile plastic tissue culture flasks of 25 cm^2 surface area (Costar^R N. 3055, Cambridge). Evolution of parasite transformation was made each five days by examining a drop of medium fixed in methanol and stained in Giemsa stain. Metacyclic forms were counted in a total of 200 parasites. Study of in vitro infectivity - A VERO cell line and a murine reticulum sarcoma line (J774) were cultivated originally at 37 C in Dulbecco modified medium, pH 7.2, established by Dulbecco and Freeman (1959) supplemented with 10% of heat-inactivated fetal calf serum. The second cell line having a similar behaviour to the macrophage culture cells line (Ralph et al. 1975). Cells were previously counted and centrifuged and adhesion took place onto a 12 mm diameter cover-slides placed into tissue culture flasks (25 cm^2 growth area) containing 5 ml of medium which were incubated at 37 C for 24 hr waiting for cell adhesion. Parasites were collected from an axenic culture in a metacyclic known percentage, leading a 10 parasite/cell relation. Cells were exposed for 24 hr to parasites and incubated at 37 C. At the end of the exposure period, non interiorized parasites were removed by repeated washings. Cultures were observed daily the first 3 days. Days 4, 6 and 8, we took out cover-slides that were washed with PBS, stained in Giemsa stain and put onto slides. Infection index was evaluated by counting the infected cell number and intracellular forms randomly chosen in 200 cells. All assays were made twice. In order to rule out any possibility of T. cruzi contamination of the studied samples, PCR test was carried out once more at the end of the experiments. RESULTS Data concerning the development of the parasite in mice, one month old, infected with 2.5 x 10^5 parasites of the C23 strain of T. rangeli are shown in Fig. 1. Prepatent period was of 4 days and the peak of parasitemia reached 10^6 parasites/ml at day 14 p.i. No bloodstream parasites were seen after day 30 p.i. Although every inoculated mouse was infected, no mouse infected with T. rangeli died. Figure 1: Trypanosoma rangeli. Parasitemia evolution in mice, one month aged, infected with 25 x 10^4 trypomastigotes. The histopathological study disclosed slight inflammatory infiltration foci in brain, skeletal muscle and liver sections. No one of the studied tissues showed pseudocysts (Table I). Comparative study of metacyclogenesis of the C23 strain in the three culture media was done in Fig. 2. The best data of transformation were observed in Grace's insect medium with a peak of 32% at day 40 p.i. When LIT and TAUP media were used the initial parasite development had a progressive decline since day 20 p.i. in both cases, sharing peaks of 15% and 6%, respectively. Figure 2:
comparative study of the metacyclogenesis
of Trypanosoma rangeli in three axenic media; LIT: liver infusion
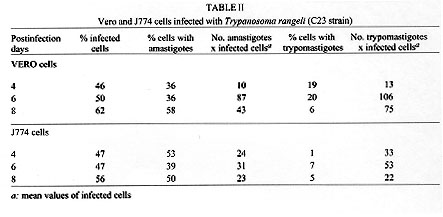
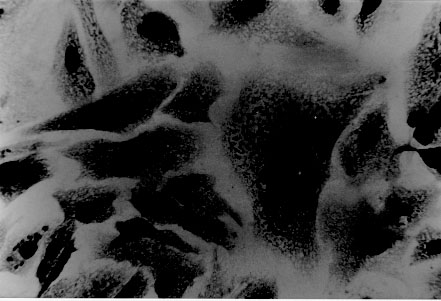
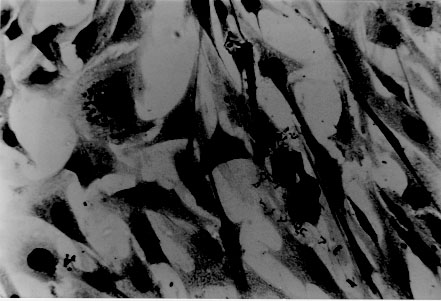
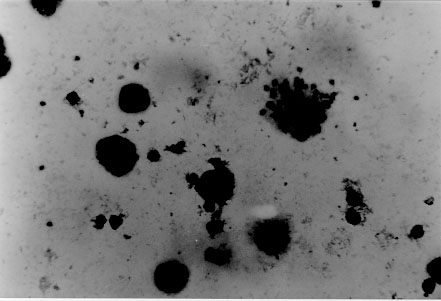
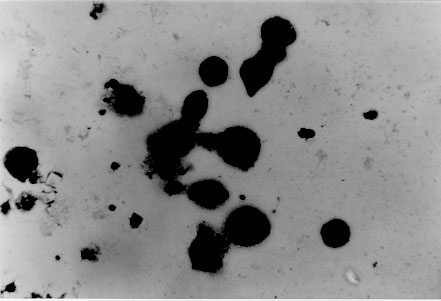
tryptose; Results of in vitro infectivity after 4 hr of incubation showed a large number of parasites adherent to cells, few of them appearing inside. At day 4 p.i. 46% of VERO cells and 47% of J774 cells showed amastigote and/or trypomastigotes forms enclosed within them. After 6 days p.i. 50% of VERO cells and 47% of J774 cells were infected, but intracellular parasites were at this time increased. Day 8 p.i. seems to show the beginning of a second cycle of infection with a large number of cells containing amastigote forms but less quantity being infected with trypomastigote forms (Table II). Figure 3, Figure 4, , Figure 5, Figure 6 and Figure 7 showed VERO and J774 cells infected by T. rangeli at different days p.i. Figure 3: VERO cells infected with Trypanosoma rangeli at day 4 postinfection. Giemsa X 400. Figure 4: VERO cells infected with Trypanosoma rangeli at day 6 postinfection. Giemsa X 400. Figure 5: J774 cells infected with Trypanosoma rangeli at day 4 postinfection. Giemsa X 400. Figure 6: J774 cells infected with Trypanosoma rangeli at day 6 postinfection. Giemsa X 400. Figure 7: J774 cells infected with Trypanosoma rangeli at day 8 postinfection. Giemsa X 200. Infection with bloodstream parasites shared no significant rate of infection. The absence of contamination of the T. rangeli utilized in this study was confirmed by PCR. DISCUSSION Important factors are decisive to get high levels of parasitemia in the experimental infection with T. rangeli other than the used strain, such as the number of trypomastigote forms contained in the inoculum. When difficulties to obtain a large number of bloodstream parasites were found, it could be interesting to have a culture medium with a good transformation rate from epimastigote to trypomastigote forms. The Grace medium showed to be the best of the studied media to obtain metatrypomastigotes (32%) in contrast with the LIT medium which shared a transformation similar with those reported for other T. rangeli strains (Urdaneta-Morales & Tejero 1985). TAUP medium was not suitable to T. rangeli growth and transformation differing from most strains of T. cruzi (Homsy et al. 1989). Metacyclic forms of C23 strain obtained from Grace culture showed to have a high infectious capacity in VERO cells and J774 cells after 4 days of incubation of parasites and cell lines together. When T. cruzi has been observed during the in vitro host-paratise interaction, varied results have been obtained: metacyclic forms of the Dm 28c clone, obtained after 72-96 hr of differentiation in an axenic medium are able to associate to 30-60% of heart muscle or fibroblastic cells within 2 hr of parasite contact and 30-50% of the associated parasites are already interiorized (Contreras et al. 1988). Indeed, authors working with bloodstream trypomasytigotes and non-professional phagocytic cells like VERO or BEMS cells (Bertelli & Brener 1980), fibroblasts (Kongtong & Inoki 1975), primary heart or skeletal muscle cells (Araujo-Jorge et al. 1986, Meirelles et al. 1986) pointed out to the need of at least 9-24 hr of parasite contact to obtain indexes varying between 1 and 9% of infected cells, depending on the parasite strain used. Comparative studies between T. rangeli and T. cruzi strains are controversial due to the heterogeneity of clonal populations within a T. cruzi strain (Deane et al. 1984, Dvorak 1984, Tibayrenc & Ayala 1988, 1991, De Diego et al. 1991, Penin et al. 1996). In spite of T. rangeli intracellular multiplication could remain in the speculative field, some authors have been able to find parasite tissular forms in vertebrate hosts (Urdaneta & Tejero 1986). Moreover, the viability of T. rangeli intracellular parasites has been suggested to be able to survive intracellularly over extended periods (Osorio et al. 1995). Few inflammatory infiltrates were found in the histopathological sections, suggesting as Osorio et al. (1995) that T. rangeli infection did not elicit a strong host response, in agreement with the low antigenicity of natural infections by T. rangeli (D'Alessandro-Bacigalupo & Saravia 1992) and the low and brief immune responses observed in experimentally infected animals (Anez et al. 1985, Urdaneta-Morales & Tejero 1986). The scarcity of intracellular forms and the possibility that parasites could be localized in lymphoreticular tissues have been argued as causes of failure to detect amastigote forms in tissular sections of the experimentally infected animals (Deane 1969, Osorio et al. 1995) as shown in our data. However, some lymphoreticular tissues have been also studied by us, such are lymph nodes. Special attention was also made to the visceral capillaries with negative results. Although it is well known that T. rangeli is difficult to establish in experimental animals (Hoare 1972) as shown by characteristically low and inconstant parasitemias, C23 strain of T. rangeli have been shown to produce high and prolonged levels of parasitemia, high infection rates in VERO and in J774 cells without serious pathological tissular findings which makes it a good model to study T. rangeli in a murine model due to its easy maintenance by mice passages. This strain could also be an interesting model to study the mixed infection by T. rangeli and T. cruzi for high, sustained and without histopathological damage. ACKNOWLEDGEMENTS To M Ayala and S Nicholls, Instituto Nacional de Salud, Bogota, Colombia for providing the C23 strain of T. rangeli. Dr Zuniga and Dr Palau benefited of BIBA Fellow, FIS and ICI Fellow, Madrid, Spain, respectively. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97094h.jpg] [oc97094d.jpg] [oc97094a.jpg] [oc97094c.jpg] [oc97094i.jpg] [oc97094f.jpg] [oc97094e.jpg] [oc97094b.jpg] [oc97094g.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}