 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ultrastructure of endogenous stages of Eimeria ninakohlyakimovae Yakimoff & Rastegaieff, 1930 Emend. Levine, 1961 in experimentally infected goat Luiz S Vieira/^+, Jose D Lima*, Mucio FB Ribeiro*, Ivete A Bozzi**, Elizabeth RS Camargos** Centro Nacional de Pesquisa de Caprinos, EMBRAPA, Fazenda TrEs Lagoas,
Caixa Postal D-10, 62011-970 Sobral, CE, Brasil Received 9 October 1996
Code Number: OC97096
File Sizes:
Text: 17.3K
Graphics: Line drawings and photographs (jpg) - 251.7K
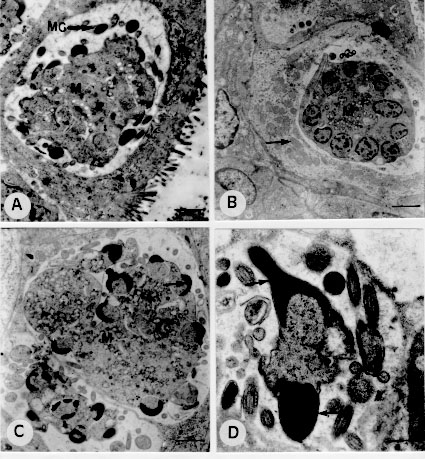
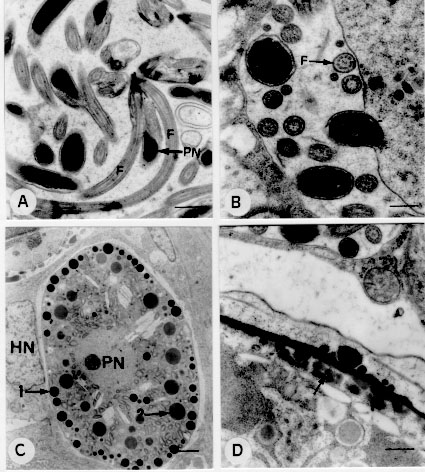
The ultrastructure of endogenous stages of Eimeria ninakohlyakimovae was observed in epithelial cells of cecum and colon crypts from a goat experimentally infected with 2.0 x 10^5 oocysts/kg. The secondary meronts developed above the nucleus of the host cell. The nucleus first divides and merozoites then form on the surface of multinucleated meronts. Free merozoites in the parasitophorous vacuole present a conoid, double membrane, one pair of rhoptries, micronemes, micropore, anterior and posterior polar ring, a nucleus with a nucleolus and peripheral chromatin. The microgamonts are located below the nucleus of the host cell and contain several nuclei at the periphery of the parasite. The microgametes consist of a body, a nucleus, three flagella and mitochondria. The macrogamonts develop below the nucleus of the host cell and have a large nucleus with a prominent nucleolus. The macrogametes contain a nucleus, wall-forming bodies of type I and type II. The young oocysts present a wall containing two layers and a sporont. Key words: goat - Eimeria ninakohlyakimovae - ultrastructure - Coccidia - Eimeria The ultrastructure of Eimeria species infecting domestic animals has been extensively studied (Scholtyseck 1973, Chobotar & Scholtyseck 1982), but the coccidia of goats have been less contemplated in studies of this nature. Light microscopy studies of the endogenous cycle of Eimeria ninakohlyakimovae in goats have been partially described in naturally (Sayin 1964, Qadir 1980) and experimentally (Balozet 1932, Norton 1986) infected animals. The ultrastructure of this species is still unknown. In the present report it is described the ultrastructure of endogenous stages of E. ninakohlyakimovae in a goat experimentally infected at two days of age. MATERIALS AND METHODS Oocysts of E. ninakohlyakimovae were obtained by the inoculation of coccidia-free kids with oocysts collected from naturally infected goats. The pure culture was selected, after serial inoculations in coccidia-free kids, using morphological and biological characteristics to separate the E. ninakohlyakimovae from other species. Oocysts collected were sporulated in 2.5% potassium dicromate solution and stored at 4 C until use. A two day old crossbred goat was inoculated orally with 2.0 x 10^5 sporulated oocysts of E. ninakolyakimovae/kg and killed 15 days after infection. Cecum and colon fragments were collected and fixed in 2.5% glutaraldehyde in 0.1M phosphate buffer (pH 7.4), and postfixed in 2% osmium tetroxide, diluted in the same buffer. The material was then dehydrated in ethyl alcohol and embedded in Epon 812. Ultrathin sections, stained with uranyl acetate (Watson 1958) and lead citrate solution (Reynolds 1963), were examined under a Zeiss EM 10 transmission electron microscope. RESULTS Development and ultrastructure of secondary meronts and merozoite formation - Secondary meronts were observed in the epithelial cells of cecum and colon crypts, where they were located above the nucleus of the host cell inside a parasitophorous vacuole. Trophozoites contained a nucleus with peripheral chromatin and electron-lucent vacuoles. Meront development started with nuclear division and merozoites develop from an elongation of a region close to each nucleus, forming the apical complex of the merozoite (Fig. 1 A). During this stage, the anterior region of the merozoite is free and the posterior region is linked to the meront. Finally, the posterior end of the merozoite is released from the meront (Fig. 1B). Figure 1: electron-micrographs of second generation merogony and of merozoites of Eimeria ninakohlyakimovae in the large intestine epithelial cells of experimentally infected goat, 15 days after infection. A: division and migration of meront nuclei (arrow), scale bar = 2.1 um; host cell nucleus (HN). B: formation and liberation (arrows) of merozoites, scale bar = 0.8 um; residual body (RB). C: merozoite, scale bar = 0,4 um; conoid (C), rhoptrie (R). D: merozoite, scale bar = 0.3 um; micronemes (MI), micropore (MP), elecron-dense bodies (CED). Merozoite ultrastructure - Merozoites, bounded by a double membrane, were detected free inside parasitophorous vacuoles. They contain a nucleus with a nucleolus and peripheral chromatin. The apical complex consists of the conoid, a pair of rhoptries and several micronemes ( Figs 1C, 1D). The ducts of the rhoptries run towards the anterior end and extend as far as the conoid and the posterior region, which is wider and club-shaped, extends in a caudal direction as far as approximately half the length of the merozoite (Figs 1C, 1D). The anterior polar ring was not easily observed, but the posterior ring was clearly visible. The micropore is located above the nucleus in the anterior region (Fig. 1D). Two to four spherical electron-dense bodies are usually located in the anterior region (Fig. 1D). Microgamont ultrastructure and microgamete formation - The microgamonts inside the parasitophorous vacuoles developed below the nucleus of epithelial cells of the cecum and colon crypts (Fig. 2 A). They were round or slightly oval in shape and were bounded by a membrane. Immature microgamonts contained 2 to 19 nuclei distributed through the cytoplasm or localized at the periphery of the parasite, with a nucleolus and peripheral chromatin (Fig. 2B). The microgametes develop in two phases: nuclear division occurs first and then the microgametes differentiate. After nuclear division, the nuclei migrate toward the periphery of the parasite and chromatin concentrates in one half of the nucleus (Fig. 2C). The nucleus presents two regions, an electron-dense one that develops toward the parasite membrane and a less dense one that remains attached to the microgamont (Fig. 2D). After maturation, the posterior portion of the microgamete is released from the micro-gamont. Figure 2: electron-micrographs of microgamonts of Eimeria ninakohlyakimovae in the large intestine epithelial cells of experimentally infected goat, 15 days after infection. A: microgametogenesis localization; microgamont (M) with microgametes (MG) situated below the host cell nucleus (HN), scale bar = 1.6 um. B: microgamont in process of nuclear division, scale bar = 2.1 um; host cell mitochondria (arrow). C: microgamont (M), scale bar = 1.6 um; chromatin (arrow). D: nuclei migration (arrows) of microgamont to formation of microgametes; scale bar = 0.6 um. Microgamete ultrastructure - The microgametes were detected free inside parasitophorous vacuoles. They are elongated or slightly curved in shape, and cross-sections are rounded. They consist of a body associated with an elongated mitochondrion and an electron-dense nucleus bounded by two membranes. They have two flagella originating from the anterior end which are longer than the microgamete (Fig. 3 A). In cross-sections of the flagella, nine pairs of microtubules localized at the periphery were observed, as well as a central pair (Fig. 3B). Figure 3: electron-micrographs of microgamete, macrogamete and oocyst formation of Eimeria ninakohlyakimovae in the large intestine epithelial cells of experimentally infected goat, 15 days after infection. A: microgamete, scale bar = 0.6 um; flagellum (F), parasite nucleus (PN). B: transversal section of flagellum (F), scale bar = 0.4 um. C: macrogamete, scale bar = 2.5 um; parasite nucleus (PN), wall-forming body type I (1), wall-forming body type II (2), host cell nucleus (HN). D: formation of the oocyst wall, scale bar = 0.4 um; coalescence of WFB II (arrow). Development and ultrastructure of macrogametes and oocysts - Macrogamonts were detected below the nucleus of epithelial cells of cecum and colon crypts. They are ovoidal in shape and are located inside a parasitophorous vacuole bounded by only one membrane. They have a large nucleus, with a prominent nucleolus. The wall-forming bodies of type II (WFB II) are the first to be formed; they contain a scarcely electron-dense structure with a spongy aspect and are usually close to organelles resembling rough endoplasmic reticulum. Wall-forming bodies of the type I (WFB I) were observed only in more advanced stages. They are electron-dense, spherical and smaller than WFB II. In macrogametes, WFB I are arranged in a row at the periphery of the parasite and WFB II are concentrated in the central part (Fig. 3C). Young oocysts (Fig. 3D) are bounded by a double wall, formed by coalescence of WFB I (outer) and WFB II (inner). DISCUSSION The ultrastructural development of secondary meronts and the formation of merozoites of E. ninakolyakimovae are similar to those of E. bovis (Sheffield & Hammond 1967, Dubremetz & Elsner 1979), E. ovinoidalis (Kelley & Hammond 1973) and E. praecox (Lee & Millard 1971a). The electron-lucent vacuoles detected in the trophozoites of E. ninakolyakimovae are comparable to those of E. acervulina, described by Michael (1975) as alimentary vacuoles. The ultrastructure of E. ninakolyakimovae merozoites is similar to that of other Eimeria species (Sheffield & Hammond 1966, Kelley & Hammond 1973, Fernando 1974). The dense bodies observed in the anterior region of E. ninakohlyakimovae merozoites are comparable to those of E. acervulina, described by Michael (1975) as protein granules. The microgametogenesis of E. nina-kolyakimovae, characterized by nucleus multiplication and by microgamete differentiation, is similar to the general pattern reported for other Eimeria species (Scholtyseck et al. 1972, Ball et al. 1988). The microgametes of E. ninakolyakimovae have the same structure as the microgametes of other Eimeria species (Scholtyseck et al. 1972, Ball & Pittilo 1988, Ball et al. 1988). The ultrastructural aspects of macrogamete development and oocyst wall formation of E. ninakolyakimovae generally present the same sequence as described for other Eimeria species infecting mammals (Scholtyseck et al. 1971) and birds (Scholtyseck et al. 1971, Michael 1975, Wheat et al. 1976, Elwasila 1984, Pittilo & Ball 1984). The WFB I and WFB II observed in the macrogametes of E. ninakolyakimovae are shared by most Eimeria species (Scholtyseck et al. 1971). The WFB II of E. ninakolyakimovae are formed before the WFB I, as also occurs for other Eimeria species (Lee & Millard 1971a,b, Scholtyseck et al. 1971), except E. truncata (Gajadhar & Stockdale 1986), in which WFB I are formed before WFB II. The size of WFB I and WFB II varies among the different Eimeria species. The WFB I of E. ninakolyakimovae are smaller than the WFB II, as is the case for E. bovis and E. auburnensis (Scholtyseck et al. 1966, 1971), but differ from those of E. perforans, E. tenella and E. maxima (Scholtyseck et al. 1971, Elwasila 1984). REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97096b.jpg] [oc97096c.jpg] [oc97096a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}