 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
RESEARCH NOTE Detection of sylvatic dark morphs of Triatoma infestans in the Bolivian Chaco Francois Noireau/^+ , Rosemary Flores*, Teresa Gutierrez**, Jean Pierre Dujardin*** Institut Francais de Recherche Scientifique pour le Developpement en
Cooperation (ORSTOM), CP 9214, La Paz, Bolivia ^+Corresponding author. Fax: 591-2-39 14 16. E-mail: noireauf@bisnet.tfnet.org Received 22 April 1997; Accepted 5 June 1997
Code Number:OC97108
Sizes of Files:
Text: 9.9K
Graphics: Line drawings and photographs (jpg) - 10.1K
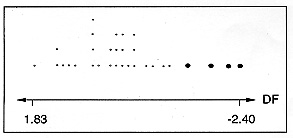
Key words: Reduviidae - Triatoma infestans - sylvatic focus - Bolivia Triatoma infestans, the most important vector of Chagas disease in the southern part of Latin America, is considered to originate from the Cochabamba valley in Bolivia because this is the only region where true sylvatic foci have been confirmed (RA Torrico 1946 An Lab Central Cochabamba 1: 19-23, JP Dujardin et al. 1987 J Med Entomol 24: 40-45, H Bermudez et al. 1993 Am J Trop Med Hyg 49 suppl.: 371). Genetic arguments fully support this assumption (JP Dujardin et al. 1997 Med Vet Entomol, in press). Except for this region where sylvatic colonies of T. infestans were found amongst rock-piles associated with wild guinea-pigs (Galea musteloides), T. infestans is said to be confined to domestic and peridomestic habitats throughout its distribution area (RL Usinger et al. 1966 An Rev Entomol 11: 309-330, CJ Schofield 1988 Biosystematics of Haematophagous Insects ed. MW Service 37: 284-312, Clarendon Press, Oxford). Nevertheless, former reports indicate that T. infestans specimens were occasionally found in sylvatic conditions in areas of Argentina (S Mazza 1943 Prensa Med Arg 30: 1-23, JFR Bejarano 1967 2 Jorn Entomoepidemiol Arg 3: 171-196), Paraguay (CJ Velasquez & G Gonzalez 1959 Rev Goiana Med 5: 357-373) and Brazil (MP Barretto et al. 1963 Rev Inst Med Trop Sao Paulo 5: 289-293). In a review dedicated to such records, Bejarano (1967 loc.cit.) mentions adults, nymphs and eggs collected in a great variety of ecotopes: under rocks or trunks of fallen trees, in hollow trees or under bark, in shelters or burrows of marsupials and rodents (Galea sp., Microcavia sp.), in bird-nests occupied by owl (Strix sp.) or by small rodents (Graomys sp.). Observations of flight by T. infestans attracted by light were also reported by this same author in Argentina. These data were completed by JA Cichero et al. (1984 Chagas 1: 33-37) who collected sylvatic T. infestans in bird-nests inhabited by Furnariidae and in burrows of Calomys callosus. Some of these reports could be due to misidentifications, and it is now generally admitted that T. infestans does not maintain sylvatic foci in these areas because most specimens were found in ecotopes relatively close to human dwellings (Usinger et al. loc. cit.). Although T. infestans is not considered a good flyer (CJ Schofield & JNS Matthews 1985 J Trop Med Hyg 88: 211-222), it can disperse in nature over distances of at least 2 km (N Schweigmann et al. 1988 Med Vet Entomol 2: 401-404). Recent observation has drawn attention to the existence of a possible T. infestans sylvatic focus in the Bolivian Gran Chaco. During the dry season of 1995 and 1996, we performed light-collections of Triatominae in an isolated site of this semi-arid region covered by a dry deciduous forest (Tita camp, way to Izozog). The nearest human dwelling was located beyond 30 km. The light system consisted of a vertical white cloth illuminated by the association of one 12-V fluorescent black light tube and one 150-V mercury vapor light. The light was operated for three nights (6 and 7 September 1995, 1 October 1996). A total of 78 starved triatomine bugs were collected between 6:30 and 9:30 p.m. and identified by morphology and genetics (isoenzyme analysis) as T. guasayana (45 specimens) or T. sordida (31). In addition, two adult specimens of undetermined taxonomic status were collected. Morphologically, they were very similar to domestic T. infestans collected in the Bolivian Chaco, except for their overall darker coloration with small yellow markings on the connexivum. They were also closely related to T. melanosoma, a completely black triatomine collected in peridomestic bird-nests in the Province of Misiones, Argentina (H Lent et al. 1994 Mem Inst Oswaldo Cruz 89: 353-358). In October 1996, a total of 11 parrot-nests located at a height of 10-15 m from the ground were studied in a radius of 10 km around the Tita camp. They were all inhabited by Aratinga acuticaudata (Psittacidae). Eight nests were positive for triatomines (73%). Four nests contained T. platensis and T. sordida. Three nests were occupied by T. platensis only. In the last one, two adult specimens of the same undetermined status were found in association with T. platensis. All the collected dark morphs (two males and two females) were compared with T. infestans by morphometrics and isoenzyme electrophoresis. Morphometrics of the head was performed according to JP Dujardin et al. (1997 Acta Trop, in press) and metric characters compared with 27 domestic T. infestans collected in Izozog, the nearest area of endemicity. Multivariate analysis showed evident differences between the two populations (Fig.). To attempt to estimate the level of relatedness among T. infestans from Izozog and these dark morphs, enzyme electrophoresis was carried out on cellulose acetate membranes, according to procedures previously described by JP Dujardin and M Tibayrenc (1985 Ann Soc Belge Med Trop 65: 271-280). Data were obtained from 53 Triatominae (49 T. infestans from Izozog and the four dark morphs) at 21 gene loci (Gpd-1, Gpd-2, Dia-1, Dia-2, Dia-3, Got, Gpi, Idh-1, Idh-2, Lap-1, Lap-2, Mdh-1, Mdh-2, Me-1, Me-2, Pep-1, Pep-2, Pep-3, Pep-4, Pgm and 6-Pgdh). No diagnostic locus was found to distinguish the dark morphs from T. infestans. Figure : Dotplots (P Sasieni & P Royston 1996 Appl Stat 45: 219-234) showing the distribution along the discriminant function (DF) of four sylvatic dark morphs (bold dots) and 27 domestic Triatoma infestans (small dots), the latter collected from two villages of the same area (Izozog). Each dot represents a single individual. Multivariate discriminant analysis was performed on log-transformed data of four head non-redundant measurements (outer eyes distance, distance between ocelli, length of the antenniferous tubercle and total head length). Discrimination was highly significant (P < 0.0001) even after removing size variation (P < 0.007; detailed results not shown). This finding opens new hypothesis about the geographical expansion of both sylvatic T. infestans or T. melanosoma, and could bring some insight into the taxonomic relationship between these two species. The isolated nature of collecting sites, the color and metric differences but the isoenzyme similarity of our four specimens with domestic T. infestans are good arguments for inferring a separate population of the same species occurring in this sylvatic area. The repeated collections (Sept. 1995 and Oct. 1996) are a further argument to support this hypothesis. Nevertheless, such a wild focus should be confirmed by collecting nymphs and eggs in the natural habitats. Besides the hypothesis of a new wild focus of T. infestans, we cannot discard the other hypothesis of incompletely black sylvatic morphs of T. melanosoma occurring in the Bolivian Chaco. Indeed, except for its coloration, T. melanosoma is morphologically similar to T. infestans. T. melanosoma has been discovered in northeastern Argentina and initially considered as a subspecies of T. infestans (A Martinez et al. 1987 Chagas 4: 479-480). Recently, it was elevated to a new species status by Lent et al. (loc. cit.). However, like the four dark morphs from the Chaco, T. melanosoma does not show any difference by isoenzyme analysis from domestic T. infestans (JP Dujardin, unpublished data). According to this second hypothesis, the spread of T. melanosoma would now include Bolivia. Lastly, this finding of dark morphs could help in understanding of evolutionary relationship between T. infestans and related species such as T. melanosoma and T. platensis (Usinger et al. loc. cit., J Pereira et al. 1996 Heredity 77: 47-54). This project was funded by ORSTOM, MAE of France and AVINA Foundation of Switzerland. Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97108a.jpg] |
| |||||||||

{kind=link}