 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Role of protein kinases in antigen-activation of peripheral blood mononuclear cells of Schistosoma mansoni infected individuals Celia A Almeida, Alfredo M Goes^+ Departamento de Bioquimica e Imunologia, Instituto de Ciencias Biologicas, Universidade Federal de Minas Gerais, Caixa Postal 486, 30161-970 Belo Horizonte, MG, Brasil ^+Corresponding author. Fax: +55-31-441.5963. E-mail: goes@mono.icb.ufmg.br Received 16 April 1997; Accepted 30 June 1997
Code Number:OC97115
Sizes of Files:
Text: 22.3K
Graphics: Line drawings and photographs (jpg) - 121.4K
T cell recognition of antigens displayed on the surface of antigen presenting cell results in rapid activation of protein tyrosine kinases and kinase C. This process leads to second messengers, such as inositol phosphates and diacylgycerol, and phosphorylation of multiple proteins. The role of different protein kinases in the activation of peripheral blood mononuclear cells (PBMC) from Schistosoma mansoni infected individuals was evaluated using genistein and H-7, specific inhibitors of protein tyrosine kinase and kinase C, respectively. Our results showed that proliferation in response to soluble egg antigen or adult worm antigen preparation of S. mansoni was reduced when PBMC were cultured in presence of protein kinase inhibitors. Using these inhibitors on in vitro granuloma reaction, we also observed a marked reduction of granuloma index. Taken together, our results suggest that S. mansoni antigen activation of PBMC involves protein kinases activity. Key words: schistosomiasis - T cell activation - protein tyrosine kinase - protein kinase C The helminth Schistosoma mansoni is responsible for the disease schistosomiasis characterized by a granulomatous inflammation, developed around parasite eggs trapped in host tissues (Warren et al. 1967, Boros & Warren 1970). Large granulomas result as a consequence of the host's T cell sensitization to antigens from the eggs. A feature of the immunobiology of schistosomiasis is the gradual and spontaneous reduction in the size of granulomatous inflammation around the continuously incoming eggs. This phenomenon, termed immunomodulation (Andrade & Warren 1964) has been the subject of numerous studies that have implicated the participation of cellular as well as subcellular mechanisms in regulation of granulomatous process (Ottensen & Poindexter 1980, Goes et al. 1991, Cheever et al. 1992). The transmembrane signalling mechanisms triggered by S. mansoni antigens and the biochemical changes in response to T cell receptor (TCR) activation are not completely understood. It is currently accepted that the signal transduction cascade is initiated by activation of protein tyrosine kinases (PTK) (Samelson et al. 1986, June et al. 1990, Chan et al. 1994), which results in the phosphorylation of a number of cellular proteins, such as phospholipase C-g1 (PLC) (Weiss et al. 1991). Activated PLC, in turn, catalyzes the hydrolysis of membrane phosphoinositides to generate inositol 1,4,5-trisphosphate (IP3) and diacylglycerol, second messengers responsible for the mobilization of cytoplasmic free calcium (Ca^2+) and activation of protein kinase C (PKC), respectively (Imboden et al. 1985). Although it is conceivable that protein kinases are involved in activation of peripheral blood mononuclear cells (PBMC) from chronic schistosomiasis patients, direct proof is not yet available. In order to verify if protein kinases play an essencial role in the regulation of granulomatous hypersensitivity to S. mansoni antigens, we studied whether protein kinase inhibition could interfere with proliferation and in vitro granuloma reaction. We evaluated the effects of the specific PTK inhibitor, Genistein (Ogawara et al. 1989) and PKC inhibitor, 1-(5-Isoquinolinysulfonyl)-2-methylpiperazine dihydrochloride (H-7) (Hidaka et al. 1984) in activation of PBMC from chronic schistosomiasis patients stimulated with S. mansoni antigens. MATERIALS AND METHODS Study population - Chronic intestinal schistosomiasis patients were selected based on clinical and parasitological stool examination for the presence of S. mansoni eggs (Hoffman et al. 1934) in Fundacao Nacional de Saude, Santa Luzia, State of Minas Gerais, Brazil. The patient protocols used throughout this study were approved by a human subject ethics committee in Brazil. Protein kinase inhibitors - Genistein was used as PTK-selective (Ogawara et al. 1989) and H-7 as PKC-selective inhibitor (Hidaka et al. 1984). They were purchased from Sigma Chemical Co. (St. Louis, MO) and dissolved in DMSO at 10 mM stock solution. Cell preparations - PBMC were isolated from heparinized blood by Ficoll-diatrizoate density gradient centrifugation (LSM, Organon Teknika, Charleston, SC) as previously described (Goes et al. 1991). These cells were cultured in RPMI medium containing 1.6% L-glutamine, 300 U/ml of penicillin, 0.3 mg/ml of streptomycin, 0.05 mg/ml of gentamicin and 10% heat inactivated AB+ human serum (RPMI 10% AB) in 96 well flat-bottomed tissue culture plate. Every experimental group was maintained at 37 C in a 5% CO2 incubator. Cell proliferation assay - PBMC from chronic schistosomiasis patients were cultured in 96 well flat-bottomed plates in RPMI 10% AB at the concentration of 3 x 10^5 cells/well in a final volume of 200 ul. Genistein or H-7 were added to each well at the same time as S. mansoni antigens. Cultures were stimulated with 25 ug/ml of soluble egg antigen (SEA) or soluble worm antigen preparation (SWAP). As controls, cells were cultured without inhibitor and/or schistosoma antigens. The cultures were set up in triplicate and incubated for 120 hr. For the last 18 hr of incubation, cells were pulsed with 0.5 uCi/well of [methyl-^3H]thymidine (ICN Biomedicals Inc., Irvine, CA). Cells were then harvested by an automated cell harvester and the incorporated radioactivity measured by a liquid scintillation beta counter, was expressed as counts per minute (cpm). Percentage of inhibition of proliferation was calculated as described by Chiaffarino et al. (1994):
inhibition (%) =
100 -{cpm (Antigen + inhibitor) - cpm (control + inhibitor)}x 100
-----------------------------------------------------------------
cpm (Antigen) - cpm (control)
In vitro granuloma reaction - Two hundred S. mansoni-conjugated polyacrylamide beads were distributed in a volume of 50 ul/well in 96 well flat-bottomed culture plates (Doughty et al. 1987). Cells at the concentration of 3 x 10^5/well in a final volume of 200 ul RPMI 10% AB were cultured in the presence and absence of genistein or H-7. Every experimental and control group was set up in triplicate and maintained at 37 C in a 5% CO2 incubator. In vitro granuloma reaction was evaluated on day 5 after culture initiation by quantification of cellular reactivity around the beads using a phase contrast inverted tissue culture microscope. Cellular reactivity was determined by morphological observation based on the following criteria: the number of cells binding to the beads; visual evidence of blast transforming cells accompanied by cellular migration; and adherent cell layers surrounding the beads. A total of 300 separated determinations of cellular reactivity were made for each experimental group. A numerical score equivalent to the following classification was assigned to each cell-bead reaction observed: (1) no cells binding to the bead; (2) < 5 cells binding to the bead; (3) =/>5 cells binding to the bead; (4) =/> 5 cells binding to the bead accompanied by a circumoval mononuclear cell migration and blast transformation; (5) adherent cell layer attached to the bead accompanied by circumoval mononuclear cell migration; (6) multiple cell layers surrounding the bead accompanied by mononuclear cell migration. The total score was then summed and the resultant mean expressed as the granuloma index (GI) +/- standard deviation (SD). Granulomatous reactivity to PB-antigen (PB-SEA or PB-SWAP) was compared to the nonspecific binding of PBMC to polyacrylamide beads not conjugated to antigen (PB). Percentage of inhibition of in vitro granuloma reaction was calculated as follows:
inhibition (%) =
100 -{ GI (PB-antigen + inhibitor) - GI (PB + inhibitor)} x 100
---------------------------------------------------------------
GI (PB-antigen) - GI (PB)
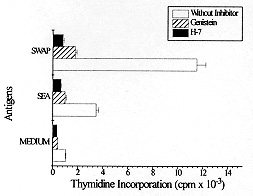
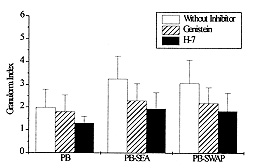
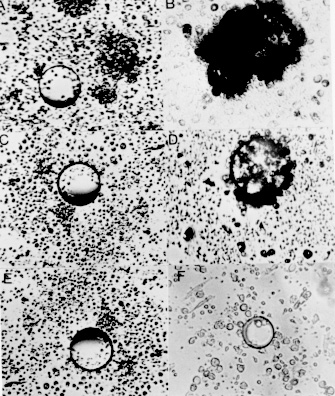
RESULTS Given the involvement of protein kinases in T cell activation, we initially investigated the effect of selective inhibitors genistein and H-7 on PBMC proliferation induced by S. mansoni antigens measuring [^3H] thymidine incorporation. We cultured PBMC from chronic schistosomiasis patients with 50 mM of genistein or with 55 mM of H-7. The data in Fig. 1 demonstrate that PBMC response to schistosomal antigens was effectively impaired by inhibitors. These results were obtained when the inhibitors were present during the entire assay. The proliferation of PBMC cultured with SEA or SWAP in presence of genistein or H-7 was significantly smaller than that observed in the absence of inhibitors. When PBMC were assayed for the ability to proliferate in response to SEA, it was found that genistein and H-7 reduced thymidine uptake by approximately 70% and 80%, respectively. The thymidine uptake of PBMC cultured with SWAP was inhibited by 80% in presence of genistein and 90% in presence of H-7. Fig. 1: effect of protein kinase inhibitors on PBMC proliferation by Schistosoma mansoni antigens. 3 x 10^5 cells/well from schistosomiasis patients were cultured in 96 well flat-bottomed with or without 50 uM of genistein or 55 uM of H-7 for five days. Proliferative responses were determined by pulsing the cells with 0.5 uCi/well [^3H] thymidine for the last 18 hr of culture. Results represent means +/- Standard Error of at least six experiments. PBMC from patients with chronic intestinal schistosomiasis mansoni were tested for in vitro granuloma reaction to SEA-conjugated beads (PB-SEA) and to SWAP-conjugated beads (PB-SWAP) as can be seen in Fig. 2. The cellular reactivity to PB-SEA and PB-SWAP was statistically greater as compared to PB alone (P<0.05). We observed inhibition of granulomatous reaction to PB-SEA (Fig. 3) and PB-SWAP when PBMC were cultured with protein kinase inhibitors. The reactivity of cells to PB-SEA was reduced by about 60% when PBMC were cultured with genistein and 50% with H-7. The granuloma index diminished 65% and 50% when PBMC were cultured with PB-SWAP in presence of genistein and H-7, respectively. Fig. 2: effect of protein kinase inhibitors on in vitro granuloma reaction. 3 x 10^5 cells/well were incubated in 96 well flat-bottomed plate in the presence or absence of 50 uM genistein or 55 uM H-7. These cells were cultured for five days with polyacrylamide beads (PB) or beads conjugated to SEA (PB-SEA) or SWAP (PB-SWAP). The granuloma index was evaluated by cellular reactivity around the beads. Results represents means +/- Standard Deviation of at least nine experiments. Fig. 3: morphologic illustration of in vitro granuloma formation. A total of 3 x 10^5 PBMC from chronic schistosomiasis patients were cultured with PB (A) and PB-SEA (B) without protein kinase inhibitors, with PB (C) and PB-SEA (D) in the presence of genistein and with PB (E) and PB-SEA (F) in the presence of H-7. DISCUSSION Engagement of the TCR with antigen or anti-TCR monoclonal antibody induces at least three rapid biochemical changes that are of probable significance in signal transduction: an increase in the level of free intracellular calcium (Imboden et al. 1985), the activation of protein kinase C (Chen et al. 1986, Nel et al. 1986), and an increase in tyrosine phosphorylation (Campbell & Sefton 1990, June et al. 1990, Lane et al. 1990). Increased protein phosphorylation is central and essencial to the stimulation of cell proliferation by myriad factors, for example epidermal growth factor (Ushiro & Cohen 1980) and colony stimulating factor 1 (Scherr et al. 1985), each of which binds to a receptor that is a tyrosine protein kinase. Protein phosphorylation is also induced by many cytokines, including interleukin 2 (Saltzman et al. 1988). Therefore the possibility that it plays an important role in lymphocyte activation cannot be ignored. However, much remains to be learned of the complex mechanism involved in antigen activation of PBMC. In our study we decided to investigate if protein kinases are involved in activation exerted by both soluble and conjugated S. mansoni antigens. Specific inhibitors of protein kinases provide useful means to examine it. The extensive use of genistein (Mustelin et al. 1990, Chiaffarino et al. 1994) and H-7 (Yamamoto et al. 1986, Nel et al. 1987) in different cellular systems prove that these drugs inhibit most, if not all, protein kinase-mediated biological responses. Then we also analyzed the participation of PTKs and PKC in PBMC activation induced by SEA and SWAP through the use of specific inhibitors. We found that genistein and H-7 caused an inhibition of proliferation of PBMC from chronic schistosomiasis patients and that 50 and 55 mM respectively, were inhibitory, without affecting the PBMC viability as measured by trypan blue dye exclusion (data not shown). Hence the observed effect should be due to inhibition of protein kinases in S. mansoni induced activation. It should be noted that thymidine uptake in lymphocytes stimulated with SEA and SWAP in presence of H-7 was lower than thymidine uptake when lymphocytes were stimulated in presence of genistein. In fact, H-7 is a competitive inhibitor of ATP and therefore inhibits other kinases with variable potencies. Protein kinase inhibitors were also employed to verify if protein kinases are necessary for in vitro granuloma reaction. The results obtained showed a significant reduction in granuloma index when PBMC were cultivated with genistein and H-7. The observed reduction in proliferation and on in vitro granuloma reaction when PBMC were cultivated with protein kinase inhibitors suggests that during immunomodulation, protein kinases might be under control, down-regulating the immune response to S. mansoni antigens. In conclusion, our results indicate the involvement of protein kinases in the activation of PBMC induced by SEA and SWAP. Further effort should be made to understand the signalling mechanisms involved in activation and immunomodulation of schistosomiasis. This research was supported by grants from the Conselho Nacional de Pesquisa (CNPq), Pro-Reitoria de Pesquisa da Universidade Federal de Minas Gerais (PRPq-UFMG) and from Fundacao de Amparo a Pesquisa de Minas Gerais (FAPEMIG). REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97115b.jpg] [oc97115c.jpg] [oc97115a.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}