 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
RESEARCH NOTE Characterization of Rotavirus Strains with Unusual Electrophoretic Profiles Joana D'arc P Mascarenhas, Carlos F Arias*, Luis Padilla-Noriega*, Suzana Lopez*, Rosa Helena P Gusmao, Yvone B Gabbay, Alexandre C Linhares/^+ Instituto Evandro Chagas, Fundacao Nacional de Saude, Av. Almirante
Barroso 492, 66090-000 Belem, PA, Brasil Received 27 January 1997; Accepted 25 June 1997
Code Number:OC97144
Size of files:
Text: 16.5K
Graphics: Tables (jpg) - 18.6K
Line drawings and photographs (jpg) - 27.8K
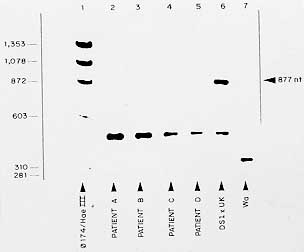
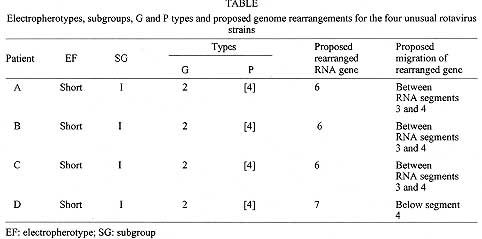
Key words: rotavirus - unusual - electrophoretic profiles Group A rotaviruses are currently recognized as the most important cause of acute gastroenteritis among both infants and young children worldwide, accounting for about one million deaths per year in developing countries (I DeZoysa et al. 1985 Bull WHO 63: 569-583, NR Blacklow & HB Greenberg 1991 N Engl J Med 325: 252-264). Rotaviruses are members of the family Reoviridae and their genome consists of 11 discrete segments of double-stranded (ds) RNA surrounded by a triple-layered shell virion (MK Estes & J Cohen 1989 Microbiol Rev 53: 410-449). The usual RNA electrophoretic profile of group A rotaviruses displays four different size classes containing RNA segments 1 to 4, 5 and 6, 7 to 9 and 10 and 11. Those rotavirus strains with genomic patterns not matching this basic configuration may be regarded as either non-group A rotaviruses or group A rotaviruses with rearranged genome (E Mendez et al. 1992 Arch Virol 125: 331-338). In this context, avian-like rotavirus strains have been recognized which display five segments in the first dsRNA class (H Brussow et al. 1992 J Clin Microbiol 30: 67-73, RHP Gusmao et al. 1994 J Diarrhoeal Dis Res 12: 129-132). Each of the 11 RNA segments codes for a single polypeptide, with the exception of gene 11 which codes for two polypeptides. Three of the virus structural proteins (VP4, gene 4; VP6, gene 6; and VP7, genes 7, 8 or 9) induce antibodies that are used for classification of rotaviruses on serological basis (Estes & Cohen loc. cit.). VP6 forms the middle shell of the virus and induces cross-reactive group- and subgroup-specific, non-neutralizing antibodies. On the other hand, the two surface proteins VP7 and VP4 independently induce type-specific neutralizing antibodies that form the basis for the current classification of group A rotaviruses into G (VP7) and P (VP4) serotypes (Estes & Cohen loc. cit., Y Hoshino et al. 1985 Proc Ntl Acad Sci USA 82: 8701-8704). Among rotavirus strains from both human and animal origin, 14 G serotypes and at least 20 P types (and 10 P serotypes) have been described (GF Browning et al. 1991 J gen Virol 72: 1059-1064, MK Estes 1996 p. 1625-1655. In BN Fields, DM Knipe, PM Howley (eds), Rotaviruses and their replication, Raven Press, New York). In addition to serotypes, nucleotide and aminoacid sequence analysis of gene segment 4 have identified six distinct rotavirus VP4 genotypes among human rotaviruses, designated as P[8] (prototype strain Wa), P[4] (strain DS-1), P[6] (strain M37), P[9] (strain AU228), P[10] (strain 69M) and P[12] (strain PA169) (M Liu et al. 1988 Virology 163: 26-32, K Taniguchi et al. 1989 J Virol 63: 4101-4106). Most of human isolates from diarrhoeic children fall into four groups, as based on the current binary system proposed for rotavirus characterization, as follows: P[8], G1; [P8], G3; [P8], G4; and P[4], G2 (JR Gentsch et al. 1992 J Clin Microbiol 30: 1365-1373, A Steele et al. 1993 J Clin Microbiol 31: 1735-1740). Rotavirus strains characterized as being [P8], G1and [P4], G2 seem to be the most common worldwide (BK Das et al. 1994 J Clin Microbiol 32: 1820-1822, NBG Rasool et al. 1993 Arch Virol 133: 275-282). Although limited, the available data in Brazil concerning the occurrence of rotavirus G and P genotypes indicate that the predominant strains are essentially the same as those most commonly found worldwide. Thus, [P8], G1 and [P4], G2 account for more than 50% of isolates (MCS Timenetsky et al. 1994 J Clin Microbiol 32: 2622-2624, V Gouvea et al. 1994 J Clin Microbiol 32: 1408-1409, JPG Leite et al. 1996 Arch Virol 141: 2353-2364). In these studies, the detection of mixed infections is an indicator that in vivo reassortments may occur, as previously reported in northern Brazil (JDP Mascarenhas et al. 1988 Virus Res 14: 235-240). Of interest, these investigators have shown that rotavirus genotype [P8], G5 accounts for a significant proportion of cases of acute diarrhoea among infants and young children throughout the country. The routine use of rotavirus RNA electrophoresis in polyacrylamide gel has made possible the identification of "atypical" genomic patterns which may derive either from a non-group A strain or from group-A viruses that underwent rearrangements in their genome segments (SM Lipson & MH Kaplan 1988 J Diarrhoeal Dis Res 10: 97-100). The previous detection of rotavirus strains in Belem with an unusual avian-like profile (i.e. displaying five segments in the first dsRNA size class) raised the question of possible naturally occurring genomic rearrangement of human strains or, less likely, interspecies transmission (Gusmao et al. loc. cit.). The identification of two rotavirus variants containing rearrangements involving the RNA segments 5 and 7 has been reported before for human strain Wa (Mendez et al. loc. cit.). In general, it has been demonstrated that altered, higher molecular weight RNA segments are generated by rearragement of the wild type gene(s), resulting in partially duplicated sequences (M Gorziglia et al. 1989 Virology 170: 587-590, SM Matsui et al. 1990 J Virol 64: 120-124). This report deals with the characterization of unusual, avian-like rotavirus strains obtained from four children, aged less than two years, who were admitted to a public hospital in Belem, between July and September 1993. Stool specimens were collected as soon as possible following the detection of a diarrhoeal episode, as well as from control, non-diarrhoeic patient. Faeces were then placed in phosphate buffered saline (PBS), pH 7.4, for rotavirus screening by using a commercial ELISA kit (DAKOPATTS ELISA, Copenhagen, Denmark), as previously described (TH Flewett et al. 1989 Bull WHO 67: 369-374). Further subgrouping and G serotyping of rotavirus-positive strains were performed essentially as described by K Taniguchi et al. (1987 J Infect Dis 155: 1159-1166), by using monoclonal antibodies against each of the two subgroups and four human G serotypes which were kindly provided by Dr Shozo Urasawa, Department of Hygiene and Epidemiology, Sapporo Medical College, Sapporo, Japan. Extraction of dsRNA from stool samples was performed and RNA was electrophoresed through a 5% polyacrylamide slab gel using discontinous buffer system, as previously described (AJ Herring et al. 1982 J Clin Microbiol 16: 473-474). The gels were then fixed in ethanol/acetic acid, stained with silver nitrate, washed, and developed with NaOH/formalin. The VP4 (P) typing was done by polymerase chain reaction (PCR) following the method of Gentsch et al. (loc. cit.). Briefly, the dsRNA was first reverse transcribed and amplified by PCR through two successive steps in which both consensus and nested primers were used. The first amplification was performed in the presence of primers designed to amplify an 876-bp DNA fragment corresponding to a highly conserved region of gene 4 among strains belonging to VP4 genetic groups 1 to 4. Portions of this fragment were subsequently used as template in a second PCR by using, in addition to the plus - sense consensus primer, a mixture of type-specific primers designed to amplify the 267-bp (P type 3), 345-bp (type 1), 391-bp (type 4) and 483-bp (type 2) fragments of gene 4. The PCR products were then separated in a 1.5% agarose gel in TAE (Tris-base 40 mM, acetic acid 0.1%, EDTA 1 mM), and it was stained with ethidium bromide. The events of the four cases can be summarized as follows. Patient A was a non-diarrhoeic 18-months old female hospitalized on July 7, 1993; patient B was a 14-months old female admitted on July 5 and developed acute gastroenteritis on July 11 (stool sample obtained three days later); and patient C was a 3-month old infant male admitted on July 9 with acute diarrhoea which persisted for three days. This infant was kept hospitalized because of acute respiratory infection (ARI) and, four days after the first episode was resolved, a second one occurred lasting five days: faecal sample was obtained on the fourth day of diarrhoea. On September 23, 1993 a further unusual strain was detected in stools of a diarrhoeic 6-months old female infant (patient D) who was hospitalized one week before: this child was hospitalized with ARI and developed diarrhoea four days later. All four strains were found to belong to subgroup (SG) I and VP7 G serotype (ST) 2. The atypical genomic profiles of rotaviruses excreted by the four study children are shown in Fig. 1. Fig. 1a shows three identical "short" electrophoretypes (patients A, B, and C), in which the 11 RNA fragments are distributed into four distinct clusters in a 5-1-3-2 configuration. The further, also "short" electro-phoretype (patient D) displays a 5-2-2-2 distribution of the 11 RNA segments (Fig. 1b). (Electrophoresis could not be performed with all strains at the same time, as some of the specimens collected in July 1993 exhausted after several RNA extractions). Fig. 2 shows the results of P typing of the four rotavirus strains by PCR (second amplification). The VP4 gene of all four strains were amplified with specific primers for DS1 (P[4] genotype, P1B serotype), thus being associated with P[4], G2 proteins of human strains. Fig. 1: electrophoretypes of atypical rotavirus strains obtained from hospitalized children in Belem, Brazil. Fig. 2: RT-PCR typing of atypical rotavirus strains obtained from hospitalized children in Belem, Brazil. Lane 1, molecular weight markers [FX174 digested with HaeIII]. Lanes 2 to 5, products amplified from dsRNA from atypical rotavirus strains [lane 2, patient A; lane 3; patient B; lane 4, patient C; lane 5, patient D]. Lanes 6 to 7, products amplified from ds RNA from human strains: [lane 6, P4 genotype control (DS1 X UK virus); lane 7, P8 genotype control (Wa virus) (band of 877 nt in this lane represents the product of the first PCR that was re-amplified in the second PCR, because of carry over of some primers from the first reaction). The detection of unusual, avian-like rotavirus strains among hospitalized children in Belem, led us to postulate that an interspecies transmission might be occurring within hospital environment. This was originally sustained by the striking similarity between the atypical RNA patterns of rotavirus strains in the present study and that recorded for turkey rotavirus strains in previous investigations (Brussow et al. loc. cit.). The hypothesis was that patient A (the apparent index case) became infected in the community and was excreting atypical rotavirus strain when admitted to the hospital because of an acute respiratory disease (Gusmao et al. loc. cit.). The subsequent infection of patients B and C with the same strain strongly suggested that a nosocomial transmission of this atypical rotavirus strain had occurred. The further, two months later detection of an additional avian-like rotavirus strain (patient D) also suggested that cross-species transmission might be occurring, as demonstrated by a few authors throughout the world (O Nakagomi & T Nakagomi 1989 J Virol 63: 1431-1434, G Gerna et al. 1992 J Clin Microbiol 30: 9-16). The second hypothesis (proved further to be the true one) atributted the atypical RNA configuration (resembling those of avian origin) to genomic rearrangements in human rotaviruses, as previously reported (Mendez et al. loc. cit.). All four strains were confirmed to have a VP7 serotype G2, and to belong to subgroup I, as determined by using monoclonal antibodies. On the other hand, the PCR gene of all four rotavirus strains was clearly amplified with primers specific for DS1 strain, which bears P4 genotype specificity, according to the nomenclature proposed by Estes and Cohen (loc. cit.); this corresponds to VP4 (P) serotype 1B and relates to the G2VP4 proteins in human rotavirus strains. Taken together, these observations confirm the hypothesis that the four previously characterized as avian-like rotavirus strains are in fact human rotavirus strains which underwent genomic rearrangement (Y Hoshino & AZ Kapikian 1994 Trends in Microbiol 2: 242-249). It is likely that rotavirus strains obtained from patients A, B and C have undergone rearrangement by a partial duplication of RNA segment 6, which migrates more slowly (probably between RNA segments 3 and 4, if a comparison is made with RNA segments 1 to 4 of the short RNA pattern control virus), as shown in Fig. 1. Although genome rearrangements of rotaviruses have been reported by several authors, these usually involve genes coding for non-structural proteins (i.e. RNA segments 5, 8, 9, 10, and 11). Although atypical for group A, rearrangements occurring in gene 6 have been found, producing RNA profiles of great diversity (S Pedley et al. 1984 J gen Virol 65: 1141-1150, S Shen et al. 1994 J Virol 68: 1682-1688). Regarding the rotavirus strain obtained from patient D, which also bears P[4], G2, SGI and short electrophoretype specificities, it seems that the rearranged gene is RNA segment 7, that migrates below segment 4, if compared with S2 rotavirus strain control. Mendez et al. (loc. cit.) have recently characterized a gene 7 genomic rearrangement that is very similar to the strain of patient D, except that it displays a "long" electrophoretic profile. A Table summarizes the electropherotypes, subgroups, G and P types and proposed genome rearrangements for the four unusual rotavirus strains. Although originally characterized as a very unusual phenomenon, mainly identified in rare cases of immunodeficient hosts (JJ Eiden et al. 1985 Pediatr Infect Dis 4: 632-637, KT Dolan et al 1985 J Clin Microbiol 21: 753-758), genomic rearrangements have proven to occur frequently. As in the present study, these atypical strains were found to circulate for months in immunocompetent children as a nosocomial infection (TG Besselaar et al. 1986 Arch Virol 87: 327-330), besides their circulation in a variety of healthy animal hosts (ME Thouless et al. 1986 Arch Virol 89: 161-170, RB Bellinzoni et al. 1987 J Clin Microbiol 25: 952-954, TN Tanaka et al. 1988 Arch Virol 98: 253-265). Even though our strains turned out not to be of avian origin, the peculiar electrophoretic patterns they have could serve as a useful epidemiologic marker to trace both the distribution and frequency of infections due to a single rotavirus strain, particularly at hospital environment. In addition, the fact that genomic rearrangements might change the structure and function of encoded proteins (U Desselberger 1996 Adv Virus Res 46: 69-95) may have important implication concerning future strategies of rotavirus vaccines. Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97144a.jpg] [oc97144c.jpg] [oc97144b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}