 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
External Morphology of the Immature Stages of Metacuterebra apicalis Antonio Cesar Rios Leite^+, Paul Williams Departamento de Parasitologia, Instituto de Ciencias Biologicas,
Universidade Federal de Minas Gerais, Caixa Postal 486, 31270-901 Belo
Horizonte, MG, Brasil
This work was partially supported by "Conselho Nacional de Desenvolvimento Cientifico eTecnologico" of Brazil. Received 6 May 1997; Accepted 17 July 1997
Code Number:OC97145
Sizes of Files:
Text: 24K
Graphics: Line drawings and photographs (jpg) - 178.3K
The second and third instar larvae, and the pupa of Metacuterebra apicalis (Guerin-Menevilli), are described based on light and scanning electron microscope observations. Key words: Metacuterebra apicalis - Diptera - Cuterebridae - immature stage - larva - pupa - bot fly Cuterebrid bot flies are indigenous to the New World, where their larvae cause primary cutaneous myiasis in mammals, including man (Catts 1982). Metacuterebra is restricted to the Neotropical Region. In nature, the larval stages of M. apicalis parasitize rodents (Guimaraes 1971, Guimaraes et al. 1983, Bossi & Bergallo 1992, Vieira 1993). The life cycle of this species was described by Leite and Williams (1988). The morphological characters of the M. apicalis adults were partially described, under light microscope (LM) (Lutz 1917), and redescribed by LM and scanning electron microscope (SEM) observations (Leite & Williams 1989). Based on LM and SEM studies, the egg and the first instar larva of M. apicalis have been described (Leite & Williams 1989). To complement our previous studies on the immature stages of M. apicalis, the second and third instar larvae, and the pupa are described herein. MATERIALS AND METHODS Three groups, each with six white rats, were infected by introducing two, five or ten first instar larvae of M. apicalis into the conjunctiva of the eye (Leite & Williams 1989). The infected rats were kept individually in wire covered plastic cages and were given a commercial rodent food and ad libitum access to water. On the 9th and 18th days of infection, the rats were killed with ethyl ether and cutaneous lesions were removed. Second and third instar larvae were dissected from surrounding tissues. Some rodents were maintained until third instar larvae emerged from their hosts; these free-larvae were maintained until survivors transformed into pupae. Material destined for LM studies were fixed in Dietrich/San Jean fluid for 2 hr and then stored in 70% ethanol until examination. Material for SEM examination were killed in hot water (70 C), fixed in 70% ethanol, dehydrated in a ethanol series, submitted to CO2 critical point drying and spattered with gold. At least eight specimens of each immature stage were studied. RESULTS
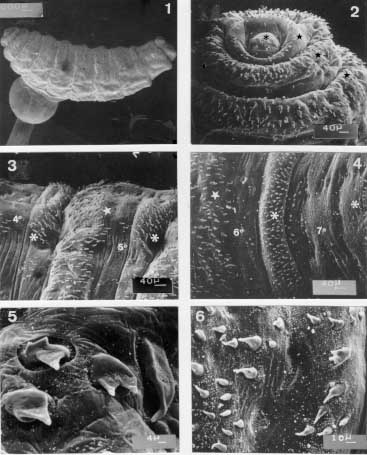
Scanning electron micrographs of the second instar larva of Metacuterebra apicalis collected on the ninth day post-infection of rats. Fig. 1: whole body in lateral view. Fig. 2: anterior end showing the pseudocephalum (asterisk), and the three thoracic segments (star) in ventral view. Fig. 3: anterior (star) and posterior (asterisk) spines of the fourth (4 ) and fifth (5 ) abdominal segments in ventral view. Fig. 4: anterior (star) and posterior (asterisk) spines of the sixth (6 ) and seventh (7 ) abdominal segments in ventral view. Fig. 5: anterior spines of the fourth abdominal segment showing spines with two and three tips (arrow). Fig. 6: anterior spine of the sixth abdominal segment. Bar = mm. Second instar larva - On the ninth day of infection, the second instar larva (Fig. 1) is subcylindrical in shape, pale in colour, and measures 5.58 - 7 mm in length and 1.24 - 1.79 mm wide at the level of the thorax. The pseudo-cephalum bears tuberiform antennae. The first thoracic segment (Fig. 2) is surrounded anteriorly by small curved thorn-like spines, each with a backwardly directed simple tip. Occasionally, bare patches were seen in the anterior region. The second and the third thoracic segments (Fig. 2) are anteriorly encircled by backwardly directed thorn-like spines with simple tips; a few spines with two- and three-pointed tips were seen on the third thoracic segment. From the first to the fifth abdominal segments (Fig. 3), anterior spines are similar to those on the third thoracic segment. The sixth abdominal segment bears anterior thorn-like spines (Fig. 4) with backwardly directed simple tips. On the seventh abdominal segment, there are a few anterior spines, all implanted on the ventral surface (Fig. 4). Posteriorly, the second to the eighth abdominal segments have spines with forwardly directed simple tips. Usually the posterior spines are larger than the anterior ones. The second abdominal segment has fewer spines than the following segments. Spines with two, three or multiple tips ( Figs 5, 6) occur in the anterior region of the second to the sixth abdominal segments and in the posterior margin from the third to the fifth abdominal segments. The opening of the anterior spiracle (Fig. 7) is in the form of an invaginated, transverse slit, and is situated lateral to the posterior margin of the second segment. The eighth abdominal segment is unarmed anteriorly.
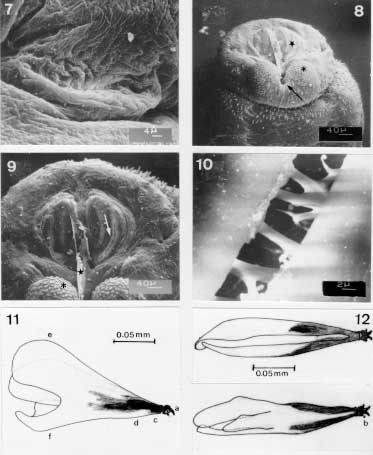
Scanning electron micrographs and camera lucida drawings of the second instar larva of Metacuterebra apicalis collected on the ninth day post-infection of rats. Fig. 7: the opening of the anterior spiracle. Fig. 8: posterior end showing the spiracular plate (star), anal tubercle (asterisk) and anus (arrow) in ventro-terminal view. Fig. 9: terminal end showing the spiracular opening (arrow), ecdisial scar (star) and anal tubercle (asterisk) in terminal view. Fig. 10: spiracular trabeculae. Fig. 11: cephalopharyngeal skeleton in lateral view. Fig. 12: idem, in dorsal (above) and ventral (below) views. Abbreviations: a: maxilla, b: dentate sclerite, c: hypophyngeal sclerite, d: tentopharyngeal sclerite, e: dorsal cornu, f: ventral cornu. Bar = mm. Both sides of the posterior end of the eighth abdominal segment (Figs 8, 9) bear a spiracular plate with an inconspicuous peritrema and two slightly curved slits, each surrounded by a brown rima, and with several trabeculae (Fig. 10). A probable ecdysis scar is visible between the spiracular plates. The anus is surrounded by two anal tubercles, each bearing small plates. The cephalopharyngeal skeleton (Figs 11, 12) is dark-brown from the anterior end to the central region, and colourless posteriorly. The maxillae are smooth, wide at the base, ventrally curved and distally pointed. The dental sclerite occupies a lateral and subterminal position on the maxilla. The hypopharyngeal and tentopharyngeal sclerite are united. The two dorsal cornua are separated distally and united basally to ventral cornua, which are fused with the pharynx on each side. The posterior margins of the dorsal and ventral cornua are markedly grooved. Third instar larva - On the 18th-day of infection, the larvae are in the third instar. This larva has a cylindrical shape, is pale in colour, measures 11.2 -18.88 mm in length and is 4.48 - 11.2 mm wide (at the level of the thorax). The pseudo-cephalum bears two tuberculate antennae, each bearing two small sensilla, and a few plates with smooth margins. The maxillae (Figs 13-15), are smooth, broad at the base, ventrally curved and sharpened distally. The three thoracic segments are each covered anteriorly by plates, which have smooth or serrated margins (Fig. 16). On each segment, the plates are imbricated and directed posteriorly in the anterior half and, thereafter, are forwardly directed. The circular spiracular aperture (Fig. 17) is well defined.
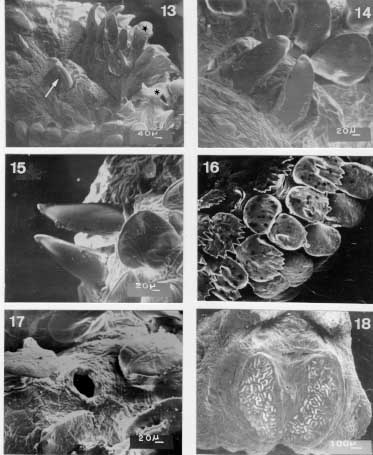
Scanning electron micrographs of the third instar larvae of Metacuterebra apicalis collected on the 18th day post-infection of rats. Fig. 13: anterior end showing the maxillae (arrow), plate spines (star), and anterior spiracular opening (asterisk) in ventro-lateral view. Fig. 14: maxillae in ventral view. Fig. 15: idem, in dorsal view. Fig. 16: plate spines on the thoracic segment. Fig. 17: anterior spiracular opening. Fig. 18: posterior spiracular plates showing the intertwined serpentine slits. Bar = mm. There are several sensilla on thoracic segments. The first seven abdominal segments have plates and sensilla, with distributions similar to those on the thorax. The eighth abdominal segment, without plates, bears light-brown thorn-like spines, which have one, two or three backwardly directed tips. Each spiracular plate is bright in colour and bears three interrupted sectors of intertwined serpentine slits (Fig. 18). Strong trabeculae are visible through the spiracular slits. The anal tubercles have small plate and the anus is anteriorly and posteriorly bordered by thorn-like spines.
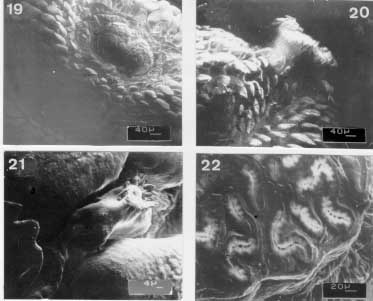
Scanning electron micrographs of the emerged third instar larva of Metacuterebra apicalis. Fig. 19: pseudocephalum in ventral view. Fig. 20: prominent anterior spiracle in ventral view. Fig. 21: cuticular sensilla. Fig. 22: posterior spiracular slits. Immediately after emerging from the host, the third instar larvae is black-brown in colour, and measures 23.94 - 35.25 mm in length by 11.7- 13.43 mm in width. The pseudocephalum (Fig. 19) is like that of the 18-day old parasitic larvae. On the tegument of the three thoracic segments there are plates, with smooth anterior edge, up to the median region; following, posteriorly, by plates with serrated anterior edge. The anterior spiracular opening has white respiratory papillae (Fig. 20). The first six abdominal segments bear plates distributed like those of the thoracic segments. The seventh abdominal segment is covered over by plates with smooth margins, whereas the eighth abdominal segment has thorn-like spines with one to three tips. The median region of the thoracic and abdominal segments have a circlet of cylindrically shaped sensilla (Fig. 21). The surface of the posterior end is similar to that of 18-day parasitic larva, such as the spiracular slits (Fig. 22).
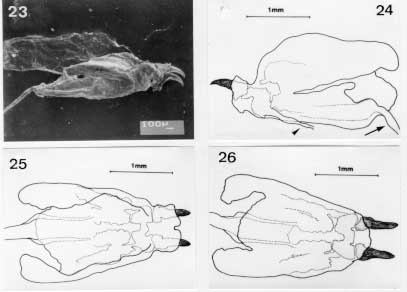
Scanning electron micrographs of the emerged third instar larva of Metacuterebra apicalis. Fig. 23: cephalopharyngeal skeleton in lateral view. Bar = mm. Fig. 24: idem, camera lucida drawing showing the salivary duct (arrow head) and oesophagum (arrow) in lateral view. Fig. 25: idem, in dorsal view. Fig. 26: idem, in ventral view. The cephalopharyngeal skeleton (Figs 23-26) of the emerged larva bears dark-brown to black maxillae, which are smooth, apically sharpened and broad basally. The dark-brown hypopharynx is fused anteriorly to the maxillae and posteriorly to the tentopharynx. The latter has a dark-brown median region, with the rest colourless. The expanded and colourless dorsal cornua, fused with colourless ventral cornua, shows a median narrow groove. The ventral cornua becomes tube-like and merges with the pharynx and, then, with the oesophagus. A small tube connects with the cephapharyngeal skeleton, anterior-laterally, is probably a salivary duct .
Light and scanning electron micrographs of the pupa of Metacuterebra apicalis. Fig. 27: pupa (above) and puparium (below) in dorsal view; x1.3. Fig. 28: anterior plate on the third abdominal segment. Fig. 29: anterior (arrow) and posterior (arrow head) plate on the fourth abdominal segment. Fig. 30: posterior plate spines on the fourth abdominal segment. Fig. 31: anterior spiracle (arrow). Fig. 32: respiratory papilla. Bar = mm. Pupa - The dark pupa (Fig. 27) is 23.13 - 27.28 mm in length and 9.83 - 13.57 mm wide, at the level of the base of the operculum. Anteriorly, the pseudocephalum and the thoracic segments are retracted posteriorly. The seventh and eighth abdominal segments are also retracted. The integument is covered with plates (Figs 28-30). Each triangular operculum has the spiracular opening situated latero-apically (Fig. 31). The Fig. 32 shows the spiracular papillae. DISCUSSION Although more than 70 species of Cuterebrids have been described (Guimaraes 1967, 1984, Catts 1982, Sabrosky 1986), little is known about their immature stages. When observed by light microscopy, the body of the second instar larva of M. apicalis is similar to those of Cuterebra species (Knipling & Brody 1940, Penner 1958, Dalmat 1942, Bennett 1955, Haas & Dicke 1958, Capelle 1970, Baird & Graham 1973), but differs from that of Dermatobia hominis (Moya Borja 1966) in general body shape and in the arrangement of the spines. The cephalonpharyngeal skeleton of M. apicalis is like those of C. buccata (Knipling & Brody 1940) and of the probable M. flaviventris (Disney 1969), except the T form of the hypopharyngeal sclerite and the flat dorsal cornu observed in C. buccata. The maxillae with a very sharp tip in C. buccata, M. flaviventris, and C. tenebrosa (Baird & Graham 1973), and thick tip in C. peromysci (Dalmat 1942) were not observed in M. apicalis. Spines with double tips seen in M. apicalis have been recorded only in second instar larva of C. tenebrosa (Baird & Graham 1973). The third instar larva of M. apicalis is unlike the third instar larva of D. hominis, by the same differences mentioned for second instar larva and by the presence of three slight curved slits in posterior spiracular plates. The external morphology of the third instar larva of M. apicalis is like that of M. schalzi (Henriksen 1942), M. chiquibulensis and M. flaviventris (Disney 1969), and of rodent Cuterebra. Rabbit Cuterebra have a single-pointed spine on cuticular plates whereas Cuterebra from rodents show multiple-pointed spines on plates (Baird 1971). The mature bot of wild primates, Allouattamyia baeri (Shannon & Greene 1926, Zeledon et al. 1957), has a typical form of Metacuterebra with dentate plates and spiracular plates with intertwined serpentine slits, but shows distinctions, such as: pronunciate furrow among the segments, as grenade-like, and plates with single and double tips. The strong maxillae in C. peromysci (Dalmat 1942) and A. baeri (Zeledon et al. 1957), and the very sharp maxillae in C. buccata and C. cuniculi (Knipling & Brody 1940), C. thomomuris (Jellison 1949), M. flaviventris (Disney 1969) and C. polita (Capelle 1970) are not observed in M. apicalis. The sensilla on M. apicalis may be like those described as papillae in C. tenebrosa larva (Baird & Graham 1973). M. apicalis, with 25 mm long, emerged from Oryzomys flavescens (Henriksen 1942) and R. norvegicus (Led et al. 1976) are between the parameters described in the present work. Similar dimensions, comparable with M. apicalis, were recorded in C. buccata (Knipling & Broady 1940), C. latifrons (Catts 1967), and C. tenebrosa (Baird 1975). The emerged larvae of C. americana (Capelle 1970) is smaller and of C. cuniculi (Haas & Dicke 1958) is larger than M. apicalis. Like other described pupae of Cuterebra [C. peromysci (Dalmat 1942) and C. tenebrosa (Baird & Graham 1973)], the M. apicalis pupa exhibits ten apparent segments, which preserve the cuticular plates of the third instar larva, and whose operculum occupies the five first tergites. The pupa of M. apicalis and Cuterebra have contractile sternites instead of contractile tergites reported in Hypoderma bovis (McLintock 1964). Puparium size of M. apicalis is similar to those of C. lepusculi (Townsend 1897) and C. tenebrosa (Baird & Graham 1973), smaller than C. cuniculi (Haas & Dicke 1958), and larger than C. peromycis (Dalmat 1942). ACKNOWLEDGEMENTS To the "Centro de Microscopia Eletronica, ICB-UFMG" for the use of the scanning electron microscope. REFERENCES Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97145c.jpg] [oc97145b.jpg] [oc97145e.jpg] [oc97145a.jpg] [oc97145f.jpg] [oc97145d.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}