 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biological Parameters and Molecular Markers of Clone CL Brener - The Reference Organism of the Trypanosoma cruzi Genome Project Bianca Zingales/^+, Maria Elizabeth S Pereira*, Katia A Almeida**, Instituto de Quimica, Universidade de Sao Paulo, Caixa Postal 26.077,
05599-970 Sao Paulo, SP, Brasil Financial support: UNDP/World Bank/WHO Special Programme for Research and Training in Tropical Diseases and FAPESP, Fundacao de Amparo a Pesquisa do Estado de Sao Paulo. Received 20 August 1997; Accepted 10 September 1997
Code Number:OC97152
Sizes of Files:
Text: 16.9K
Graphics: Tables (jpg) - 31.1K
Line drawings and photographs (jpg) - 43.1K
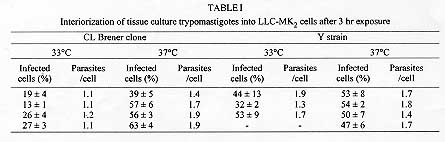
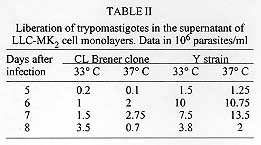
Clone CL Brener is the reference organism used in the Trypanosoma cruzi Genome Project. Some biological parameters of CL Brener were determined: (a) the doubling time of epimastigote forms cultured in liver infusion-tryptose (LIT) medium at 28 C is 58+/-13 hr; (b) differentiation of epimastigotes to metacyclic trypomastigotes is obtained by incubation in LIT-20% Grace´s medium; (c) trypomastigotes infect mammalian cultured cells and perform the complete intracellular cycle at 33 and 37 C; (d) blood forms are highly infective to mice; (e) blood forms are susceptible to nifurtimox and benznidazole. The molecular typing of CL Brener has been determined: (a) isoenzymatic profiles are characteristic of zymodeme ZB; (b) PCR amplification of a 24Sa ribosomal RNA sequence indicates it belongs to T. cruzi lineage 1; (c) schizodeme, randomly amplified polymorphic DNA (RAPD) and DNA fingerprinting analyses were performed. Key words: CL Brener - Trypanosoma cruzi - Genome Project - biological characteristics - molecular typing At difference with other Parasite Genome Projects, the participants of the Trypanosoma cruzi network present in the Trypanosomatid Genome Planning Meeting held in Rio de Janeiro in April 1994 decided to select one reference clone (CL Brener) to be used by all members. This clone derives from the CL strain which presents all important characteristics of T. cruzi: (a) it was isolated from Triatoma infestans, a strictly domiciliary vector (Brener & Chiari 1963); (b) it differentiates in liquid medium; (c) it infects cell monolayers (Alcantara & Brener 1978); (d) it has preferential parasitism of heart and muscle cells (Melo & Brener 1978); (e) it presents defined parasitemia curves and mortality in mice (Brener 1977); (f) it shows a clear acute phase in accidentally infected humans (Z Brener, personal communication); and (g) it is highly susceptible to drugs used clinically in Chagas disease (Filardi & Brener 1987). Clone CL Brener was recently isolated by Professor Zigman Brener and MES Pereira (Centro de Pesquisas Rene Rachou-Fiocruz, Belo Horizonte) from the blood of mice infected with the CL parental strain. The initial task attributed to our group was the definition of the biological characteristics of CL Brener clone as well as of molecular markers to be used to genetically characterize this organism. Furthermore, our laboratory (Instituto de Quimica, Universidade de Sao Paulo) was chosen as a Reference Laboratory for storage and distribution of the clone to members of the T. cruzi Genome Project. BIOLOGICAL AND PARASITOLOGICAL PARAMETERS OF CLONE CL BRENER Cultivation conditions and differentiation into metacyclic forms - Epimastigote forms were cultured in LIT medium containing 10% calf serum (Castellani et al. 1967), at 28 C without agitation. A doubling time of 58+/-13 hr was determined. Spontaneous differentiation of epimastigote into metacyclic forms varied from 25 to 40%. After 19 weeks of subculturing, the differentiation capacity was drastically reduced to 5-10%. Differentiation into metacyclic forms could be induced in LIT medium with different proportions of Grace's medium (20-55% v/v). After seven to ten days, 50-80% differentiation into metacyclic forms was obtained. Metacyclics could be separated from epimastigotes by chromatography in DEAE-cellulose (Whatman DE-52) columns. Differentiation in LIT:Grace medium was also reduced after prolonged subcultivation of the CL Brener clone in LIT medium. Infectivity to cultured mammalian cells - Previous studies on the morphogenesis of intracellular trypomastigotes in cell culture indicated that the amastigote-to-trypomastigote differentiation of the CL strain took place at 33 C but was inhibited at 37 C (Brener et al. 1976). This phenomenon was not observed with other strains (e.g. Y, Gilmar, G, etc.) where differentiation to intracellular trypomastigotes occurred both at 33 C and 37 C. The thermosensitivity presented by the CL strain led to the hypothesis that this strain may represent a natural "regulatory mutant" where specific gene(s) involved in differentiation have suffered mutation(s) (cf. Brener et al. 1976). We investigated whether CL Brener also presented this temperature-dependent characteristic. Monolayers of LLC-MK2 cells were exposed to a suspension of tissue culture trypomastigotes (TCT) of CL Brener employing a parasite/cell ratio of 100:1. For comparison, experiments were conducted in parallel with TCT of the Y strain of T. cruzi (Silva & Nussenzweig 1953). After 3 hr exposure at 33 or 37^ C, the percentage of infected cells and the interiorization index (parasites/cell) were determined as described (Zingales et al. 1982). Data of four independent experiments are shown in Table I. The number of TCT liberated in the culture medium of cell monolayers after five to eight days of infection (parasite/cell ratio 8:1) in cultures maintained at 33 or 37^ C was also determined (Table II). Data indicate that clone CL Brener is able to penetrate and differentiate in cultured-mammalian cells at both temperatures. In addition, the experiments suggest that the Y strain performs more efficiently the intracellular cycle when compared to CL Brener. Infectivity to mice - Parasitemia curves and mortality were determined in Balb/c mice inoculated i.p. with 5x10^5 LIT:Grace-derived metacyclic trypomastigotes of CL Brener clone and of the CL strain. Pre-patent periods of 6-7 days were observed. Data indicate that metacyclic forms of the CL-parental strain are more virulent than those of CL Brener clone. Balb/c mice were inoculated i.p. 5x10^3 blood trypomastigotes. Pre-patent periods of 6-7 days were also observed for both clone CL Brener and the CL strain. Parasitemia curves were ascending reaching 2x10^6 trypomastigotes/ml 15 days after inoculation. Mortality was 100% on day 16. Similar results were observed for the CL strain. Susceptibility to chemotherapeutic agents - Parasite sensitivity
to benznidazole and nifurtimox was assayed according to the protocol
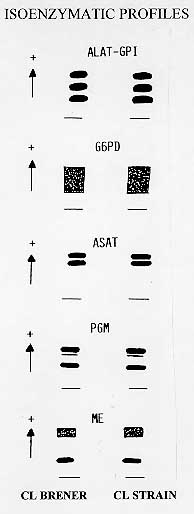
described by Filardi and Brener (1984). Two groups of MOLECULAR TYPING Isoenzymatic profiles and definition of zymodeme group - Following the protocol described by Romanha et al. (1979) isoenzymatic profiles were determined for six enzymes: alanine aminotransferase (ALAT), glucose phosphate isomerase (GPI), glucose-6-phosphate isomerase (G6PD), aspartate aminotransferase (ASAT), phosphoglucomutase (PGM) and malic enzyme (ME) and compared to standard T. cruzi zymodemes. CL Brener clone and the CL parental strain presented identical isoenzymatic profiles (Fig. 1) characteristic of zymodeme ZB (Romanha et al. 1979).
Analysis of a dimorphic sequence of the 24Sa ribosomal RNA gene - We have previously shown that the D7a divergent domain of the 24Sa ribosomal RNA gene of T. cruzi presents sequence dimorphism among parasite strains (Souto & Zingales 1993). PCR amplification of a specific region of this gene, employing oligonucleotides D71 and D72, originates fragments of 125bp (Group 1 isolates) or 110bp (Group 2 isolates) (Souto & Zingales 1993). Analogous experiments were done with the CL parental strain and CL Brener clone, originating 125bp fragments. Both products were cloned into the HincII site of M13mp19 and sequenced. Sequence alignment indicate 98% similarity of both CL Brener and CL parental strain to the 24Sa rDNA consensus sequence of Group 1-isolates (Fig. 2). The nucleotide sequence of the 125bp DNA fragment of CL Brener has been deposited in GenBank under the accession number U73959.
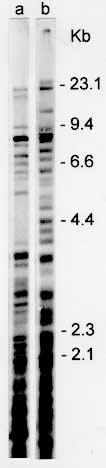
Schizodeme analysis - The technique of schizodeme analysis for genotypic characterization of trypanosomatids was introduced by Morel et al. (1980). Schizodeme analysis of CL Brener clone was done according to Goncalves et al. (1984). The patterns obtained with five restriction enzymes indicate that the schizodeme of CL Brener clone is the same as that of the CL parental strain. Data were published elsewhere (Zingales et al. 1997). Randomly amplified polymorphic DNA analysis - The basic protocol by Steindel et al. (1993) for arbitrarily primed polymerase chain reaction (AP-PCR) was followed. DNA was submitted to PCR with seven different primers including M13F, M13R, lgt11F, and lgt11R. A complex multiband profile was observed for each primer. On average 18+/-7 bands were detected, varying in intensity and size according to the primer used (Zingales et al. 1997). DNA fingerprinting - It has been shown that the human multilocus probe 33.15 (Jeffreys et al. 1985) is capable of identifying minisatellites in T. cruzi genome, generating a strain-specific complex banding pattern typical of DNA fingerprinting (Macedo et al. 1992). Genomic DNA of CL Brener was digested with BspRI or HinfI and electrophoresed in 0.8% agarose gels. Southern blots were hybridized to the 33.15 biotinylated probe. A complex multiband profile with approximately 20 bands varying in intensity and size from 2kb to 23 Kb was obtained. Bands of lower molecular weight were also detected (Fig. 3).
REFERENCES Copyright 1997 Fundacao Oswaldo Cruz The following images related to this document are available:Photo images[oc97152d.jpg] [oc97152b.jpg] [oc97152c.jpg] [oc97152a.jpg] [oc97152e.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}