 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Modulatory Role of Eosinophils in Allergic Inflammation: New Evidence for a Rather Outdated Concept
Christianne Bandeira-Melo, Renato SB Cordeiro, Patricia MR Silva,
Laboratorio de Inflamacao, Departamento de Fisiologia e Farmacodinamica,
Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ,
Brasil This work was supported by grants from CNPq and FAPERJ. Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97167
Sizes of Files:
Text: 32K
Graphics: Line drawings and photographs (jpg) - 62.9K
Tables (jpg) - 84.4K
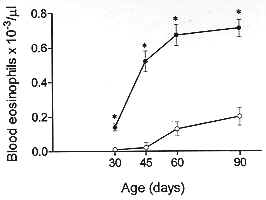
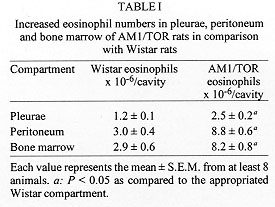
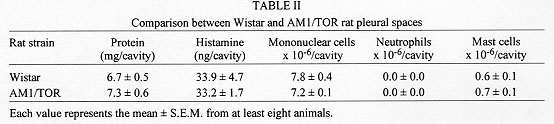
The eosinophilic response has been identified as a key alteration in the pathogenesis of asthma and other allergic diseases. A close-correlation between disease severity and eosinophilia, and the eosinophil ability to provide toxic and pro-inflammatory agents are the major elements supporting the interpretation that there is indeed a causal relationship between these phenomena. Nevertheless, controversy still persists since some studies have clearly demonstrated that eosinophil infiltration is not necessarily accompanied by tissue damage or hyperresponsiveness. In addition, there are some examples in the literature in which such alterations are not modified following abrogation of eosinophil influx. In this review it will be argued, based on a model of IgE-dependent pleurisy, that eosinophil infiltration can be associated with down-regulation of allergic inflammatory response. The potential mechanism by which eosinophils could be acting as a immunomodulatory cells in this particular system will also be assessed. Key words: eosinophil - prostaglandins - allergy - inflammation and pleurisy Eosinophils are thought to mediate inflammatory and cytotoxic events associated with allergic disorders, including bronchial asthma, rhinitis and urticaria (Gleich et al. 1993, Kroegel et al. 1994). This interpretation is in line with very well established observations in which hypereosinophilic dysfunctions are clearly associated with tissue damage, in particular endomiocardial fibrosis (Wardlaw et al. 1995). It is noteworthy that the eosinophil granule covers a spectrum of preformed highly toxic cationic proteins, including the major basic protein and eosinophil cationic protein, which are able to cause tissue damage by disrupting the membrane structure of target cells (Gleich et al. 1993). Moreover, eosinophils can also generate active substances like PAF, leukotriene C4, active species of oxygen and several cytokines which can lead to inflammation, bronchoconstriction and mucus hypersecretion, important components of the airway allergic dysfunction (Weller 1994). The pro-inflammatory effector function of eosinophils has been supported by an extensive amount of studies accumulated throughout the 1980's. One of the important observations is that the treatment of monkeys with anti-ICAM-1 monoclonal antibody clearly inhibited allergen-induced lung eosinophil influx and also prevented airway hyperresponsiveness (Wegner et al. 1990). Similar findings were obtained following treatment with an anti-IL-5 antibody (TRFK-5) (Mauser et al.1995), reinforcing the interpretation that there is indeed a causal link between eosinophilia and airway hyperreactivity. However, the evidence that another antibody to IL-5 (NC-17) (Nagai et al. 1993), or a soluble IL-5 receptor (Yamaguchi et al. 1994), suppresses allergen-induced BALF eosinophilia with little effect on bronchial hyperreactivity opposes this concept, and points out that the functional role of eosinophils in allergy is still debatle. It is interesting to note that the current pro-inflammatory profile of eosinophils replaced an opposing concept dominant until the late seventies, in which they were supposed to play an anti-inflammatory role (for an overview see Goetzl et al. 1979). According to early studies, eosinophils may be regarded as immunomodulatory cells because they can release specific enzymes able to neutralise mast cell products including histamine and PAF (Goetzl et al. 1979). The potential down-regulating role of eosinophils is also illustrated by studies showing that a significant amount of a histamine release inhibitor (further characterized as prostaglandins) is generated by human eosinophils following antigenic activation (Hubsher 1975a, b). Additional evidence for the anti-inflammatory effect of these cells comes from the finding that peritoneal neutrophil infiltration is inhibited in animals undergoing a local eosinophilia (Cook et al. 1988). The current report is an overview of our recent studies in which an inverse relationship between pleural eosinophilia and antigen-induced exudation has been documented and investigated. Association of pleural eosinophilia with down-regulation of allergic inflammatory response The pleural cavity is regarded as an interstitial tissue displaying a marked free fluid phase and a minimum fiber matrix represented by the microvilli of the mesothelial cells (Miserocchi et al. 1992). The pleural fluid and its cellular content, including eosinophils, are therefore in close contact with parietal and visceral pleurae, probably influencing and being influenced by them via secreted products. The power of this model is that the exudate can easily be removed by washing the pleural cavity with a physiologic fluid to ensure a complete analysis of inflammatory parameters, including exudate volume, protein content, total and differential leukocyte counts etc. Moreover, the remarkable ease with which the rat pleural space can be sensitised and targeted with allergen and or eosinophil chemoattractants led us to assume that such a model could be indeed useful for understanding the potential interaction between tissue eosinophil enrichment and allergic responsiveness. Experimental allergic pleurisy can be induced in rats by sensitising the animals with an intrapleural injection of a murine IgE anti-DNP monoclonal antibody, followed by intrapleural antigen challenge 24 h later (Bandeira-Melo et al. 1995, Lima et al. 1997).The response is characterized by massive mast cell degranulation and intense plasma leakage, assessed by measuring the amount of protein extravasated into the pleural cavity within 4 hr. Allergic pleurisy can also be triggered by intrapleural injection of ovalbumin into rats sensitised with a mixture of adjuvant and ovalbumin 14 days before challenge (Lima et al. 1991). In that case, exudation and mast cell degranulation are accompanied by large numbers of leucocytes in the pleural effluent. There are basically two types of experiments giving information about how an ongoing eosinophilia could affect antigen-induced pleurisy. First, the antigen challenge is performed in sensitised rats undergoing pleural eosinophilia evoked by distinct eosinophil chemoacttractants, and second, the allergic reactivity is assessed in spontaneously eosinophilic rats. Allergen-induced pleurisy in animals undergoing eosinophilia caused by eosinophil chemoactractants It has been reported that LPS failed to induce pleural plasma leakage but caused a biphasic reaction of leucocyte accumulation (Bozza et al. 1993). The first phase is marked by a neutrophilic infiltration which peaked from 3 to 6 hr and the second phase by a mononuclear cell infiltration, accompanied by a late long-lasting eosinophilia, which peaked from 24 to 48 hr, returning to the baseline level within 120 hr post-challenge. By challenging sensitised rats at different time-points before and after LPS, it is shown that the IgE-mediated pleural exudation is not modified when allergen is administered either at the beginning (neutrophil-enriched phase) or at the end of the LPS process. In contrast, exudation is significantly reduced when allergen challenge is performed 48 or 72 hr after LPS, at timings of non-selective or selective eosinophilia respectively. The results are consistent with the interpretation that during LPS-induced pleurisy the refractoriness to allergen is clearly correlated with eosinophil but not neutrophil infiltration. LPS-induced pleural eosinophilia appears to require a newly generated soluble protein, which is present in the 6 hr LPS pleural washing (LPS-PW). The transfer of a cell-free boiled sample of LPS-PW from donors to recipient rats leads to a 3-fold increase in the pleural eosinophil population within 24 hr, without modifying neutrophil or mononuclear cell counts (Bozza et al. 1993, 1994). The effect of such eosinophilia on allergic protein exudation has also been assessed, and again the presence of eosinophils is associated with a marked refractoriness to the allergen-induced exudation (Bandeira-Melo et al. 1995), giving support to the interpretation that these events are indeed inversely correlated. Correlation between eosinophil magnitude and down-regulation of allergic pleurisy is also clearly evident following PAF-mediated eosinophilia. Similar to LPS-PW, stimulation by PAF is accompanied by a selective 3-fold increase in pleural eosinophil counts 24 hr post-challenge. It is noteworthy that consecutive daily intrapleural PAF challenges lead to a progressive and selective augmentation in the number of eosinophils recovered from the pleural cavity 24 hr later (Martins et al. 1993, Silva et al. 1990). In parallel, there is a significant increase in the index of plasma leakage suppression, moving from 68% to 85% of blockade following 1 and 4 PAF stimulations respectively. The magnitude of eosinophil infiltration and allergen-evoked exudation in individual bases are shown to be inversely correlated. In contrast, exudation caused by either histamine, 5-HT or the standard mast cell degranulating agent compound 48/80 is not modified. The results indicate that the capacity of eosinophils to down-regulate exudation seems to be restricted to the context of the allergic reaction. In line with these findings, also actively sensitised rats undergoing eosinophilia evoked by either PAF or ECF-a are shown to be refractory to the allergen challenge, as evidenced by marked down-regulation of both plasma leakage and neutrophil influx (Martins et al. 1993). Allergen-induced pleurisy in animals undergoing spontaneous eosinophilia AM1/TOR rats have been studied in Brazil because they have a spontaneously systemic eosinophilia (Pimenta et al. 1987). As illustrated in Fig. 1, blood eosinophil numbers are clearly higher in AM1/TOR rats of different ages than in Wistar rats. Increased eosinophil numbers are also observed in other sites like bone marrow, pleura and peritoneum (Table I). Particularly concerning the pleural space, the eosinophilia is not accompanied by changes in neutrophil, mononuclear or mast cell numbers. In addition, no difference is observed in terms of pleural mast cell stored histamine (Table II). Nevertheless, comparing both sensitised Wistar and AM1-TOR rat, the latter is clearly less sensitive to allergen challenge (Fig. 2). It is noteworthy that this refractoriness has nothing to do with a potential difference in the production of anaphylactic antibodies, since both animals were sensitised with the same amount of IgE anti-DNP monoclonal antibody.
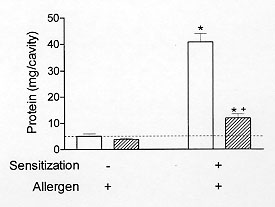
Figure 2: comparison between pleural sensitivity to allergen challenge of both sensitised Wistar (opened bars) and AM1/TOR rats (right-hatched bars). Each value represents the mean +/- S.E.M. from at least eight animals. All animals received allergen challenge. * P < 0.05 as compared to the appropriated non-sensitized group. ^+P < 0.05 as compared to sensitized Wistar rats.
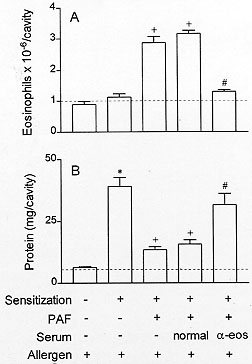
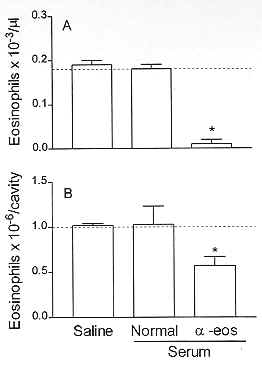
To verify whether there is a causal relationship between eosinophilia and down-regulation of allergen-induced exudation, it is important to evaluate the results of impairing the eosinophil accumulation on the suppressive phenomenon. Basically, two distinct approaches to inhibit eosinophilia have been carried out. Firstly, the eosinophil depletion obtained by means of an intravenous injection of rabbit polyclonal anti-eosinophil antiserum. Secondly, the pharmacological blockade of eosinophilia. Intravenous injection of the anti-eosinophil antiserum in naive rats leads to a selective reduction in the number of blood and pleural eosinophils 24 hr post-administration. PAF injection 24 hr before allergen challenge induces eosinophil accumulation and reduces allergic exudation. As illustrated in Fig. 3, the treatment with the anti-eosinophil antiserum inhibits eosinophilia and restores the exudatory response to allergen. Data for the effect of anti-serum treatment in naive and in sensitised animals are shown in Fig. 4 and Table III.
Figure 4: effect of anti-eosinophil antiserum treatment on blood (A) and pleural fluid (B) eosinophil numbers of naive rats. Each value represents the mean +/- S.E.M. from at least eight animals. *P < 0.05 as compared to the untreated group.
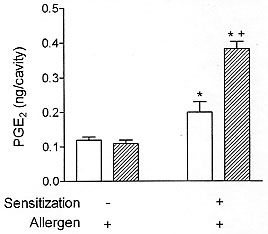
Involvement of prostaglandins in the eosinophil-mediated inhibition of allergic pleural exudation Guinea pigs and mice infected with Toxocara canis develop a brief airway hyperreactive response which is, however, followed by a marked tracheal hyporeactivity a few days later (Buijs et al. 1995a, b). Interestingly, the tracheal hyporesponsiveness, which coincides with the presence of large numbers of eosinophils in the airways, is clearly accompanied by an increased concentration of PGE2 in the bronchoalveolar lavage fluid. Moreover, according to the authors, the downward shift in airway responsiveness can be prevented by cyclo-oxygenase blockade, but not by inhibitors of lipoxygenase and superoxide formation, suggesting the involvement of prostaglandins in this suppressive phenomenon which occurs in the presence of eosinophilia. It is well established that eosinophils are able to produce PGE2 in distinct systems (Foegh et al. 1986, Giembycz et al. 1990, Kroegel & Matthys 1993). In fact, in one of the early studies approaching eosinophil function in allergy, Hubsher (1975a, b) demonstrated that immunologically-stimulated human eosinophils could inhibit in vitro histamine release through a prostaglandin-mediated mechanism. Interestingly, recent studies have provided evidence that prostaglandins of E series are indeed able to exert a negative control on the function of pivotal pro-inflammatory cell targets, including mast cells (Hogaboam et al. 1993), lymphocytes (Oppenheimer-Marks et al. 1994, Garrone et al. 1994), neutrophils (Ham et al. 1983) and macrophages (Christman et al. 1993). In line with these findings, we have verified that pleural spaces which are experiencing a PAF-induced eosinophilia and become hyporesponsive to allergen, also keep an increase in the local PGE2 concentration which, similar to the degree of allergic exudatory suppression, increased with the increasing eosinophil number (Bandeira-Melo et al. 1996). As there is no identifiable enhancement on pleural PGE2 content in naive animals undergoing PAF-related pleural eosinophilia, both ongoing eosinophilia and allergen challenge seem to be essential to evoke the PGE2 production. It is noteworthy that when Wistar and and AM1/TOR rats were compared, now in terms of anaphylactic generation of PGE2, there is indeed a significant increase in PGE2 levels in the pleural fluid of Wistar rats, but the phenomenon was clearly improved in the eosinophilic AM1/TOR rats (Fig. 5).
If the down-regulatory mechanism described herein is due to local production of PGE2, the exogenous administration of this prostanoid would be expected to have a similar suppressive effect. Indeed, orally administered misoprostol, a synthetic analogue of PGE1, attenuates plasma extravasation induced by allergen in actively sensitised animals in intensity comparable to that noted during the localised ongoing eosinophilia. Moreover, misoprostol-mediated suppression of the allergic response is not altered by indomethacin, as expected (Bandeira-Melo et al. 1996). Misoprostol has been shown to specifically bind to PGE receptors (EP receptors) and to exhibit a comparable prostaglandin pharmalogical profile (reviewed in Coleman et al. 1994), including the ability to down-regulate immunological events (reviewed in Shield 1995). We have found that PGE2 locally administered failed to alter the allergic exudatory response, which would be a consequence of its worse chemical stability and lesser resistance to the endogenous enzymatic catabolism when compared to misoprostol (reviewed in Shield 1995). It is well accepted that PGEs and their analogues increase intracellular cyclic AMP via a receptor-mediated activation of adenylate cyclase (Coleman et al. 1994). Regulation of cyclic AMP levels is also exerted by cyclic nucleotide phosphodiesterases, among which type IV isoform has been emphasised by its selectivity on cyclic AMP catabolism and sensitivity to rolipram (recently reviewed in Barnes 1995). Indeed, the combined administration of PGE2 plus rolipram significantly inhibited antigen-induced exudation, under conditions where the individual treatments are inactive. This co-operative effect between PGE2 and rolipram has been observed in other experimental models (Sinha et al. 1995) and is in line with the established interpretation that PGEs-induced inhibition of allergic response is indeed mediated by cyclic AMP. Accordingly, the intrapleural injection of the permeable analogue dibutyryl cyclic AMP significantly inhibits the allergen-induced protein exudation (Diaz et al. 1996, Bandeira-Melo et al. 1996), suggesting that an upregulation of the cyclic AMP system may indeed play a pivotal role in the hyporesponsiveness to antigen challenge expressed by the eosinophil enriched sites. Concluding Remarks We provide herein evidence that allergen-induced pleural exudation is selectively down-regulated in rats which are experiencing an ongoing eosinophilic but not a neutrophilic inflammatory response. The distinct role of eosinophils in the observed suppression was emphasized by chemical and immunological blockade of the eosinophil accumulation, which clearly restored the sensitivity to the allergen. Admitting that eosinophil infiltration and allergen-evoked exudation are inversely correlated, it is remarkable that a small increment in the former is sufficient to down-regulate the latter. This suggests that such an immunoregulatory role of eosinophils could easily take place in the microenviroment of an allergic inflammatory reaction. Furthermore, our results suggest that PGE2 seems to be implicated in the eosinophilia-related attenuation of the allergic pleurisy, in a mechanism which is probably dependent on the activation of cyclic AMP signalling pathway. Nevertheless, there is no plausible reason for thinking that eosinophils have only one function, so we should not discard the possibility that, under particular conditions, eosinophils might have their immunomodulatory activity impaired. A defect in the eosinophil ability to provide immunomodulatory mediators - like PGE2 - might be a possible explanation for the apparent duality of the eosinophil function. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97167d.jpg] [oc97167h.jpg] [oc97167e.jpg] [oc97167a.jpg] [oc97167g.jpg] [oc97167b.jpg] [oc97167c.jpg] [oc97167f.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}