 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Immune Responses of IL-5 Transgenic Mice to Parasites and Aeroallergens LA Dent, C Daly, A Geddes, J Cormie, DA Finlay, L Bignold*, P Hagan**, RME Parkhouse***, T Garate****, J Parsons*****, G Mayrhofer
Departments of Microbiology and Immunology and *Pathology, University of
Adelaide, North Tce., Adelaide, South Australia, Australia, 5005 This work was supported in part by the Australian Research Council; The Channel 7 Children's Research Fund, Adelaide, South Australia; The Medical Research Council of Great Britain. Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97168
Sizes of Files:
Text: 44K
Graphics: Line drawings and photographs (jpg) - 142.7K
Tables (jpg) - 26.6K
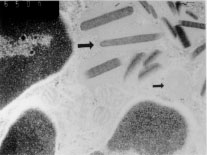
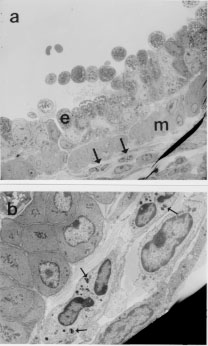
Eosinophils have long been thought to be effectors of immunity to helminths but have also been implicated in the pathogenesis of asthma. Patterns of cytokine production in the host may influence the pathogenesis of these diseases by regulating the activities of eosinophils and other components of the immune response. Mice which constitutively over-express IL-5 have profound and life-long eosinophilia in a restricted number of tissues. Although eosinophils from IL-5 transgenics are functionally competent for a number of parameters considered to be important in inflammation, untreated animals are overtly normal and free of disease. In addition, the responses of these animals when exposed to aeroallergens and helminths present a number of apparent paradoxes. Eosinophil accumulation in tissues adjacent to major airways is rapid and extensive in transgenics exposed to the aeroallergen, but even after treatment with antigen over many months these mice show no evidence of respiratory distress or pathology. Helminth-infected IL-5 transgenics and their non-transgenic littermates develop similar inflammatory responses at mucosal sites and are comparable for a number of T cell and antibody responses, but they differ considerably in their ability to clear some parasite species. The life-cycle of Nippostrongylus brasiliensis is significantly inhibited in IL-5 transgenics, but that of Toxocara canis is not. Our results also suggest that eosinophilia and/or over-expression of IL-5 may actually impair host resistance to Schistosoma mansoni and Trichinella spiralis. The pathogenesis of diseases in which eosinophils are involved may therefore be more complex than previously thought. Key words: eosinophils - interleukin-5 - helminths - asthma - allergy - Trichinella spiralis - Nippostrongylus brasiliensis - Toxocara canis Elevated levels of eosinophils are seen in the blood and tissues of animals infected with some invasive helminths, in asthmatics and some atopic individuals, in idiopathic hypereosinophilia syndrome (IHES) and in a restricted range of other diseases. Eosinophils may be a prominent feature at sites of inflammation in these diseases, though other leukocytes may also be present in large numbers. Eosinophils would seem to be well equipped to provide protection against helminths. They are capable of antibody-dependent cytotoxicity for a number of targets in vitro including the helminths Schistosoma mansoni and Trichinella spiralis (Butterworth 1984). Eosinophils are thought to damage parasites by releasing in their immediate vicinity an array of pre-formed toxic granule proteins including major basic protein (MBP), eosinophil peroxidase and eosinophil cationic protein. They may also rapidly generate and release toxic or pro-inflammatory products such as platelet activating factor (PAF) and leukotrienes. Eosinophil-mediated toxicity extends to mammalian target cells in vitro and possibly also to cultured tissues such as sections of trachea (Frigas et al. 1991). It is widely believed that asthma may in part be caused by the mis-directed or aberrant actions of eosinophils recruited to the airways by chronic exposure to antigen. To further investigate the roles of eosinophils in parasitic infections and in allergic lung diseases we have made extensive use of interleukin-5 transgenic mice (Dent et al. 1990) originally developed at the National Institute for Medical Research (NIMR, Mill Hill, UK) in the laboratory of Colin Sanderson. Our data are in some cases quite surprising and not in keeping with widely held views of the functions or eosinophils in disease. General Biology of IL-5 Transgenics The CBA/Ca IL-5 transgenics used in these studies were generated using a transgene which imparts constitutive expression of IL-5 to CD2+ cells, most of which are T cells (Dent et al. 1990). Expression of IL-5 could be very significantly enhanced by stimulation of T cells in vitro with concanavalin A and in vivo by infection with the helminth Mesocestoides corti (Strath et al. 1992), suggesting that the transgene is at least partially regulated by genomic sequences native to IL-5 and included in the transgene construct. These animals show life-long eosinophilia in the blood and in a restricted range of tissues (Dent et al. 1990). Apart from splenomegaly, even transgenics with very high transgene copy numbers are overtly normal, and in particular, show no signs of respiratory distress. B cell numbers and antibody profiles were not markedly different from those seen in non-transgenic littermates (Sanderson et al. 1994, Dent & Sanderson, unpublished results). One notable aberration which was detected early in our studies relates to breeding success. Three of the four founder transgenic lines [Tg(0IL5)C1 (Tg1), Tg2 and Tg3 with approximately 8, 49 and 44 transgene copies respectively] have been maintained as heterozygotes for more than six years without major problems. Litter sizes are comparable to those seen in nontransgenic animals of the same strain but maintained under specific-pathogen-free conditions (mean litter size +/- standard deviation for the third litters of Tg1, Tg2, Tg3 lines and a comparable nontransgenic colony were 5.7 +/- 1.8; 6.0 +/- 1.8; 6.5 +/- 1.5 and 5.5 +/- 1.91 respectively with n = 10 - 17). Despite our long-term success in breeding heterozygotes, we have not been able to successfully breed any but the lowest transgene copy line (Tg1) as homozygotes and even then the frequency of homozygotes generated is lower than expected. The reasons for this have yet be elucidated, but given that eosinophil numbers in the rodent uterus change dramatically during oestrus (Tchernitchin et al. 1974), and also the association of MBP with pregnancy in humans (Oxvig et al. 1993), excessive eosinophilia may influence the outcome of pregnancies at any of several stages. Functional Characteristics of Eosinophils from IL-5 Transgenics Chronic eosinophilia in humans with IHES has been associated with eosinophil dysfunction and ultrastructural abnormalities (Tai & Spry 1976, Henderson et al. 1988, Sokol et al. 1988). We addressed the possibility that IL-5 transgenics might not spontaneously develop respiratory disease when kept in conventional conditions because of abnormalities in many of the eosinophils which they carry. Much of this has recently been described (Dent et al. 1997). Eosinophils from IL-5 transgenics have many eosinophilic granules and at least as many as those found in eosinophils recovered from untreated or parasite-infected nontransgenic littermates (Dent et al. 1997). Many but not all of these granules have one or more electron dense bodies at the core, an ultrastructural feature typical of eosinophil granules (Fig. 1).
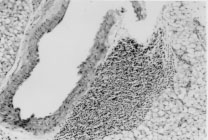
Eosinophils from IL-5 transgenics also respond in a dose-dependent manner to PAF in in vitro chemotaxis assays (Dent et al. 1997). However the real efficiency of recruitment of eosinophils from IL-5 transgenics can best be illustrated in vivo. Twenty four hours after a short exposure to aerosolized ovalbumin, large perivascular leukocytic infiltrates are evident in the lungs of immunized IL-5 transgenics ( Fig. 2). The majority of these cells are eosinophils and infiltrates are much more substantial than those in similarly treated non-transgenic littermates. In time and with further antigenic challenges the eosinophils will migrate in large numbers to occupy the submucosa of the airways.
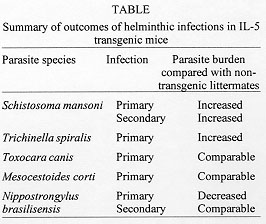
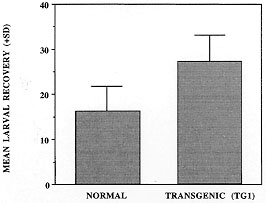
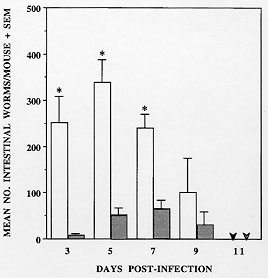
Helminth Infections in IL-5 Transgenics IL-5 transgenic mice have been experimentally infected with a number of helminth species in our laboratories. In each case parasite burdens in heterozygous transgenic mice have been compared to those in their non-transgenic littermates. We have found that over-expression of IL-5 and/or constitutive eosinophilia may enhance resistance to some but not all parasite species. However, much to our surprise, it seems that in some cases these components of the anti-helminth response may actually be disadavantageous (summarized in Table). Our first surprise came from studies conducted at NIMR with the trematode S. mansoni. Whilst some data now suggest that IL-5 and eosinophils are not important in mice for immunity to this helminth (Sher et al. 1990a,b), at the time that our experiments were begun, the only published data suggested that eosinophils may be a significant effector cell population (Butterworth et al. 1975, Mahmoud et al. 1975). We have shown that IL-5 transgenic animals have higher numbers of liver-stage S. mansoni larvae than their non-transgenic counterparts in both primary and secondary infections (Dent et al. 1997). Although vaccination appears to enhance clearance in transgenic animals, multiple vaccinations do not alter the overall effect of enhanced parasite survival in transgenic mice. S. mansoni would therefore appear either to derive some advantage from over-expression of IL-5 and/or eosinophilia, or alternatively, such a bias in the immune response in some way impairs other more protective components. In support of this outcome, we have also observed that, relative to their non-transgenic littermates, larval burdens were also higher in transgenic mice given a primary infection of the nematode T. spiralis (Fig. 3). However this is not a feature common to all nematode infections since Toxocara canis fared neither better nor worse in IL-5 transgenic mice and larval migration patterns were unaltered (Parsons, Dent et al. unpublished results). Although it has proven difficult to accurately quantitate parasite numbers in animals injected ip. with the cestode M. corti, the resulting infections follow the same protracted, and ultimately fatal course in IL-5 transgenic mice as they do in non-transgenic littermates (Strath et al. 1992).
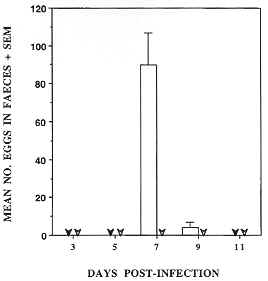
Figure 5: Fig. 5: production of eggs by Nippostrongylus brasiliensis infecting Tg2 IL-5 transgenic mice is negligible. Three faecal pellets were collected from each of three mice/group and eggs were concentrated by flotation on saturated NaCl solution and counted using a McMasters slide chamber. Values represent mean eggs/faecal pellet for each group. In this experiment eggs were only recovered from non-transgenic mice (open bars). Arrowheads represent nil eggs detected for NTg (closed) and Tg2 (open). Mice were infected by subcutaneous injection with 500 L3-stage larvae. The animals tested correspond to those depicted in Fig. 4.
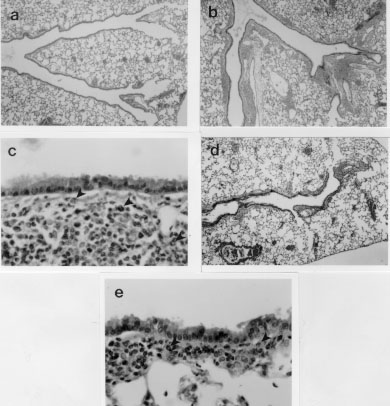
Our results with N. brasiliensis are more in keeping with the commonly perceived role of eosinophils as effectors of anti-parasite immunity. They are also in concordance with results from experiments in which IL-5 depletion led to enhancement of infections with Strongyloides venezuelensis (Korenaga et al. 1991) or Angiostrongylus cantonensis (Yoshimura et al. 1994). In this context, it may be worth noting that both of these parasites are at least distantly related to N. brasiliensis. So why should some parasites be detrimentally affected by eosinophils and others not? Further, why should infections with some parasite species appear to be worse in IL-5 transgenic mice? The answers to both questions almost certainly relate to the biology and adaptions of individual parasite species. N. brasiliensis normally produces eggs and leaves (or is expelled from) the host within ten days of infection. Changes in the number of eosinophil precursors present in the bone marrow of parasite-infected mice are not detected until approximately seven days after initiation of the infection (Strath & Sanderson 1986). By the time new eosinophils are generated in the bone marrow in response to the parasite, it has already mated and begun to produce eggs. Whilst some eosinophils may be recruited from the tissue pool before this time, either they may not be available in large numbers or they may not be able to localize to the parasite as it moves rapidly through host tissues and organs. This parasite has not been forced to live in the presence of large numbers of eosinophils and may therefore not have evolved such that it has defences which can adequately counter this cell. Consequently, when confronted with large numbers of eosinophils soon after injection into IL-5 transgenic hosts, many of the larvae may be rapidly overwhelmed. On the other hand, parasites such as S. mansoni, T. spiralis and T. canis all spend periods of months and even years in the host. It would therefore be important for their survival that they can efficiently counteract host defences of many kinds, including eosinophils. Indeed, many examples of immune evasion mechanisms adopted by helminths have been documented. More specifically, a broad range of microbes have recently been shown to utilize cytokine pathways in a variety of ways to subvert host defences. S. mansoni has apparently taken this one step further by utilizing tumour necrosis factor-a as a fecundity-enhancing factor (Amiri et al. 1992). It therefore seems possible that long-term tissue-dwelling parasites might not only evade eosinophil-mediated defences, but could also utilize IL-5 and/or eosinophil-derived factors for survival. This would seem to be the ultimate in parasitism. Responses of IL-5 Transgenic Mice to an Aeroallergen Asthma is characterized by (i) airways hyperresponsiveness (AHR), (ii) inflammation in which eosinophils are often a feature and (iii) tissue damage, which may include loss of airways epithelium, deposition of sub-epithelial collagen and plugging of the airways. Eosinophils are present in large numbers in the lungs of untreated IL-5 transgenic mice, but most are to be found in the alveolar capillaries (Dent et al. 1990) and they may also be found in smaller numbers associated with lymphoid nodules of the upper airways. We found no other symptoms of asthma-like disease in IL-5 transgenic mice kept under conventional conditions. As shown above ( Fig. 1), even small doses of ovalbumin can rapidly induce eosinophilia in the lungs of transgenic mice, but again without evidence of tissue damage. Various protocols have been used to induce tissue damage in these IL-5 transgenics, both in our own laboratory and by others (Lefort et al. 1996). We have found that tissue eosinophilia is maintained throughout three months of regular exposure to allergen (5 min exposure to a 1% OVA in saline aerosol, twice weekly). Eosinophils made up the majority of cells occupying the subepithelial tissues and some partially degranulated cells could be detected by electron microscopy in the immediate vicinity of the smooth muscle adjacent to major airways (Fig. 6a, b). However no tissue damage was detected in any of the transgenic or non-transgenic animals sampled at fortnightly intervals throughout these long-term exposure experiments.
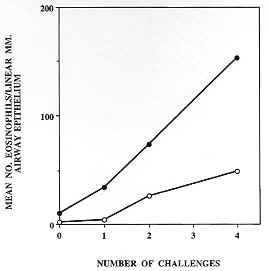
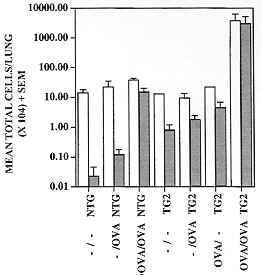
Figure 8: the number of eosinophils infiltrating sub-epithelial bronchiolar tissues of Tg2 IL-5 transgenic mice (closed circles) and their wild-type counterparts (open circles) after immunization and intensive challenge with OVA (protocol as described in Fig. 7) increases with the number of challenges, and possibly also with time elapsed after the initial challenge. Eosinophils were counted for each of two animals/group/timepoint in ten separate 1 mm. linear segments of bronchiole to a depth of 100 mm from the epithelial basement membrane. Data presented are means of mean sectional counts for each animal. A similar pattern was evident in both bronchi and trachea (data not shown). Tissue sections analyzed were similar to those shown in Fig. 7 and areas were defined using a calibrated eyepiece graticle. Figure 9: recovery of leukocytes by bronchoalveolar lavage (BAL) from the airways of mice challenged with aerosols containing OVA or saline (protocols as described in Fig. 7). BAL was performed 24 hr after the last aerosol challenge. Immunization and challenge with OVA (OVA/OVA) resulted in the recruitment of large numbers of leukocytes into the airways (open bars), the majority of which were eosinophils (closed bars) in both Tg2 IL-5 transgenics (TG2) and non-transgenic (NTG) littermates. Immunization alone (OVA/-) or OVA aerosol challenge alone (-/OVA) had minimal effect on the cellular composition of the airways relative to untreated (-/-) animals. The number of leukocytes recovered from NTG OVA/OVA mice was double that from untreated or OVA/- NTG groups and most of this change could be accounted for by an increase in eosinophils. Approximately one hundred-fold more eosinophils were recruited in transgenic than in non-transgenic animals (note log10 scale) and yet similarly treated animals (TG2 and NTG) did not develop any evidence of airways damage (see Fig. 7). BAL was performed with 3 x 1ml volumes of PBS delivered and recovered with a syringe and modified needle. Cells were enumerated with a haemocytometer and differentiated using Cytospin (Shandon) preparations stained with Giemsa (200 cells/slide), 3 mice/group.
Conclusions In studying the roles of eosinophils in both immunity to pathogens and in allergic disease, we have been forced to re-assess the current dogma on a number of issues. Our own data and that of others suggest that eosinophils may protect the host against some but not all helminth species. Our own results also suggest that the presence of high levels of IL-5 and/or eosinophilia in some way impedes destruction of at least two species of invasive helminths which both typically induce the production of eosinophils. Parasites which dwell in the host for long periods of time are likely to have evolved evasion mechanisms which negate the actions of eosinophils. They may also have taken parasitism a step further such that they utilize components of the immune response for their own benefit - this may include either IL-5 or eosinophil products. When considering the role of eosinophils in asthma, perhaps we might gain from the following analogy. If we were to examine the scene of a fire from a single snapshot, it is likely that we would be wrong in assuming that the fellows with the hoses attached to the red trucks were responsible for initiating the blaze. Eosinophils are often found in the lungs of asthmatics and indeed there is a substantial body of experimental evidence which suggests that under some conditions they may contribute to the pathogenesis of the disease. However much of the evidence is circumstantial. The logic often flows as follows: eosinophils can cause damage and they are present in large numbers where damage is manifested, therefore they are responsible for the damage. Firemen can light fires as well as put them out, and sometimes they do just that. However this is not usually a suitable explanation of the events captured in our snapshot of the conflagration. In our experimental studies of the roles of eosinophils in asthma we have established that even very spectacular eosinophilia is not sufficient to lead to allergen-induced lung disease. In the future we should focus on the as yet unknown genetically determined characteristics which regulate the ways in which eosinophils might contribute in asthma. The interplay between IL-5, eosinophils and other factors which can potentiate disease is likely to be complex and so many avenues for effective therapy of this disease have yet to be explored. Our two-pronged approach to the study of eosinophils has again focused our thoughts on the likelihood that eosinophils cannot simply be classified as end-stage effector cells or killers of microbes which occasionally misbehave and end up damaging the host unnecessarily. We must seriously consider the many products that eosinophils can secrete both in terms of their effects on microbial targets and the surrounding tissues, but also on other cells of the immune system. We must address the possibilities that there are subsets of eosinophils with different functions or that eosinophils may respond very differently depending upon the stimuli received. Eosinophils may be highly efficient killers which are also capable of contributing to tissue damage, but they may also play a role in tissue repair and even in immunoregulation. Cells found at sites of inflammation are not necessarily always contributing in a positive way to inflammation and it is possible that under some circumstances eosinophils or at least a subpopulation of these cells may negatively regulate inflammation. In the case of asthma, this could be beneficial to the host whereas for some parasitic infections, or for the mixed infections often encountered in the "real world", this may be either advantageous or detrimental. Acknowledgements To Dr Mike Worms, Mrs Ann Hallett, Ms Trudy Zimmerman and Mr Hans Schopp for technical assistance, to Drs Marilyn Henderson for helpful advice and for performing electron microscopy and Mr Dale Cavill for assistance with photography. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97168j.jpg] [oc97168h.jpg] [oc97168g.jpg] [oc97168f.jpg] [oc97168e.jpg] [oc97168c.jpg] [oc97168d.jpg] [oc97168i.jpg] [oc97168a.jpg] [oc97168b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}