 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
The Role of Interleukin5 (IL5 ) in Vivo: Studies with IL5 Deficient Mice Klaus I Matthaei^+, Paul S Foster, Ian G Young
Division of Biochemistry and Molecular Biology, John Curtin School of
Medical Research, Australian National University, P.O. Box 334, Canberra,
ACT, 0200, Australia Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97170
Sizes of Files:
Text: 26.5K
Graphics: Line drawings and photographs (jpg) - 34.3K
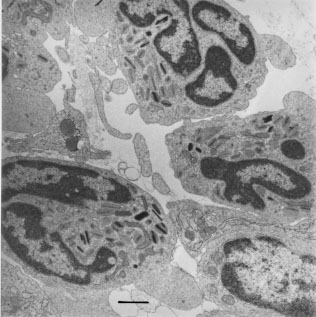
Eosinophil recruitment is a characteristic feature of a number of pathological conditions and was the topic of the recent International Symposium on allergic inflammation, asthma, parasitic and infectious diseases (Rio de Janeiro, June 3-5, 1996). Since interleukin5 (IL5) is believed to regulate the growth, differentiation and activation of eosinophils (Coffman et al. 1989, Sanderson 1992), the role of eosinophils and IL5 are closely linked. Although IL5 specifically regulates eosinophilia in vivo and this is its most well established activity, it is becoming clear that IL5 also has other biological effects. The recent derivation of an IL5 deficient mouse (Kopf et al. 1996), provides a model for exploring not only the role of IL5 and eosinophils but also other novel activities of IL5. Of note is that although the IL5 deficient mice cannot elicit a pronounced eosinophilia in response to inflammatory stimulation following aeroallergen challenge or parasite infection they still produce basal levels of eosinophils that appear to be morphologically and functionally normal. However, the basal levels of eosinophils appear insufficient for normal host defence as IL5 deficiency has now been shown to compromise defence against several helminth infections. In addition, IL5 deficient mice appear to have functional deficiencies in B-1 B lymphocytes and in IgA production. Key words: interleukin5 - knockout - eosinophils - parasite infection - B-1 B lymphocytes - IgA production Generation of IL-5^-/- mice A role for IL5 in the physiology of reproduction has been suggested since eosinophils comprise a major proportion of the abundant leukocyte populations present in the uterus. Oestrogen-regulated fluctuations in uterine eosinophil recruitment occur during the oestrous cycle and eosinophils further increase in number during the earliest stages of pregnancy, but diminish once implantation of the embryo is initiated (Robertson et al. 1994). Their possible functions in uterine tissue remodelling or in the immune response to pregnancy have not been defined. The effect of IL-5 deficiency on the fertility and fecundity of C57BL/6 mice has now been studied in some detail (SA Robertson, IG Young & KI Matthaei, unpublished). Despite a dramatic reduction in the numbers of uterine eosinophils in IL-5 deficient females mated with males of the same genotype (<15% of control mice), the numbers and viability of embryos, as well as the weights of foetuses and placentae were indistinguishable from those in control mice on day 17 of pregnancy. These data clearly show that optimal pregnancy outcome can be achieved, at least in syngeneic matings, in the absence of IL-5 and when eosinophils are severely diminished in number. The effect of IL-5 deficiency on the immune deviation required to accommodate allogeneic pregnancy is currently being investigated. Parasite infection Eosinophilia is a distinctive feature of helminth infections. Eosinophils have been postulated to be protective to the host since eosinophils arrive at the site of infection in vast numbers, and can kill some parasites in vitro (Butterworth 1984). However, evidence for a protective role in vivo has been difficult to obtain. Studies with neutralising antibodies against IL5 have suggested that eosinophils play little role in host defence (Herndon & Kayes 1992). As expected, the IL-5 deficient mice did not develop blood or overt tissue eosinophilia when infected with parasites (Kopf et al. 1996, Takamoto et al. 1997) but the effect of IL5 and eosinophil deficiency on parasite burden varied with the parasite used. The worm burden after infection with Mesocestoides corti (Kopf et al. 1996) or Toxocara canis (Takamoto et al. 1997) was not affected by IL5 deficiency. However, recent studies have shown that the absence of IL5 and eosinophils resulted in higher parasite burdens when the IL5^-/- animals were infected with more natural parasites for the mouse such as Strongyloides ratti (Ovington et al. in preparation) or Heligmosomoides polygyrus (Ovington et al. unpublished). It is possible that neutralising antibodies to IL5 may not totally ablate the actions of this cytokine. IL-5 depletion using monoclonal antibodies followed by Trichinella spiralis infection did not result in an increased parasite burden (Herndon & Kayes 1992). However, more recent studies using IL5 deficient mice infected with T. spiralis demonstrate distinctive effects due to the loss of this cytokine (Vallence et al. in preparation). Although there was only a moderate delay in the expulsion of the parasite in a primary infection, the IL5 knockout mice were significantly affected during challenge infections, typically having worm burdens twice that of their wild-type controls (Vallence et al. in preparation). Moreover, intestinal hypercontractility, an adaptive response by the host to enteric infection, was found to be moderately impaired in mice lacking IL5, suggesting a role for the eosinophil in the regulation of enteric muscle function during inflammation. Taken together these results indicate that previous studies using neutralising antibodies may not have resulted in the complete ablation of IL5, and therefore the role of IL5 during parasite infections may need to be re-examined using IL5 deficient mice. Secondly they demonstrate that in some parasitic infections IL5 contributes to host defence against parasites and this may need to be considered in connection with asthma therapy based on IL5 as a target. Since eosinophilia is seen in athymic mice infected with Toxocara canis (Takamoto et al. 1995) and since eosinophil production in bone marrow cultures in vitro can be stimulated with parasite secretions (Milbourne & Howell 1993) it has been postulated that parasites themselves could produce a molecule that is recognised by the host to be similar to IL5 (Milbourne & Howell 1993). However, infection of IL5 deficient mice with T. canis did not result in the massive eosinophilia that was seen in the control mice (Takamoto et al. 1997) indicating that host produced IL5 is responsible for the eosinophilia produced by infection with this parasite. Asthma The most striking role of IL5 and eosinophils has been shown in a mouse model of asthma (Foster et al. 1996). Eosinophils are a predominant feature of late-phase asthmatic responses and the presence of this leukocyte in the airways often correlates with disease severity and the degree of airways reactivity to spasmogenic stimuli. Sensitisation and aeroallergen-challenge of IL5^-/- mice failed to induce eosinophilia, lung damage and airways hyperreactivity indicating a central role for IL5 in the pathogenesis of allergic airways disease. The specificity of this cytokine in the inflammatory responses was demonstrated since the pathology could be totally restored by infection of the aeroallergen-challenged IL5^-/- mice with recombinant vaccinia viruses engineered to express IL5 (Foster et al. 1996). These investigations implicate the eosinophil as the primary pro-inflammatory cell involved in the development of allergic lung disease and indicates that IL5 plays a central role in the pathogenesis of asthma thereby making it an important therapeutic target for treatment of this disease (see also Foster et al. this issue). Graft rejection Eosinophils are a common infiltrating cell in rejecting pig pancreatic proislet tissue xenografts transplanted under the kidney capsule of mice, suggesting an involvement of the eosinophil in the rejection process (Simeonovic et al. 1990, Morris et al. 1995). However, pig proislet xenografts were rejected at identical rates in IL5 deficient mice even though the numbers of infiltrating eosinophils were dramatically reduced in these grafts when compared to those transplanted to normal mice (Simeonovic et al. 1997). The eosinophil appears therefore not to play an obligatory role in the rejection process. Ulcerative colitis Eosinophil recruitment into the lesions involved in inflammatory bowel disease is a characteristic of this disease (Walsh & Gaginella 1991). However, in a dextran-sulphate induced model of colitis, a reduced eosinophil infiltration into the colon was seen in IL5 deficient mice but the disease progressed normally (Stevceva et al. in preparation). These findings suggest that, in this model, eosinophils do not play a critical role in the disease process of experimental ulcerative colitis. Eosinophilia in the absence of IL-5 Of particular interest was the finding that basal levels of eosinophils were present in the IL5 deficient mice (Kopf et al. 1996). It is now clear that the residual eosinophils, although greatly reduced in numbers, are able to migrate to the sites of parasite infection (Kopf et al. 1996, Takamoto et al. 1997), xenograft rejection (Simeonovic et al. 1997), and to sites of ulcerative colitis (Stevceva et al. in preparation) or to the uterus after oestradiol administration (Kopf et al. 1996). Morphologically the eosinophils in IL5 deficient mice are normal; eosinophilic granules with normal crystalline inclusions are seen under the electron microscope in eosinophils that have migrated to the site of M. corti infection (see Fig.). Furthermore, after infection of IL 5 deficient mice with T. canis the eosinophils appeared to degranulate in the presence of the worms since structures resembling Charcot-Leyden crystals were seen (Takamoto et al. 1997). These results taken together with the fact that a blood eosinophilia can be elicited in IL5 deficient mice by the intravenous administration of eotaxin (Mould et al. 1997), or in the peritoneal cavity with agents such as LPS, PAF and VGSE (Matthaei et al. in preparation) indicates that the residual population of eosinophils are functional in the absence of IL5. Importantly, IL5 and eotaxin use different pools of eosinophils to induce eosinophilia. The intravenous administration of IL5 to IL5^-/- or normal animals mobilises eosinophils from the bone marrow, whilst the injection of eotaxin does not (Mould et al. 1997). This suggests that in normal and IL5 deficient animals there are two pools of eosinophils that can be mobilised, one present in the bone marrow that is dependent on IL5 and another that can be mobilised by eotaxin; possibly by sequestering eosinophils which are migrating through non-inflamed tissues, into the circulation. These observations again support the observation that morphologically normal and apparently functional eosinophils can be produced in the complete absence of IL5. Whether the residual eosinophils are the same as those produced in IL5 dependent eosinophilia is currently being investigated.
IgA production In vitro studies with purified IL-5 and in vivo studies with recombinant IL-5 have revealed that this cytokine functions to enhance secretion of LPS-induced-IgA by acting as a terminal differentiation factor for secretory (s)IgA^+B cells (Takatsu et al. 1985, Coffman et al. 1987). Promotion of IgA production is thought to be regulated by IL-5 alone (Coffman et al. 1987) or synergistically with TGF-b (Sonoda et al. 1992), IL-6 (Kunimoto et al. 1989) and IL-4 (Murray et al. 1987). Reports that IL-5 can stimulate the secretion of IgM and IgG by Staphylococcal A-activated B cells (Bertolini et al. 1993), further suggests that IL-5 can enhance the secretion of immunoglobulin classes other than IgA. Recent work has investigated the effects of IL-5 in a murine model when it is expressed by a live attenuated aroA strain of Salmonella serotype Dublin (SL5631) (Whittle et al. in press). Following oral or intranasal immunisation of mice with the recombinant IL5Salmonella, enhanced IgA and IgG responses against the bacteria were found. Expression of the recombinant mIL-5 also appeared to confer some advantage to the bacteria with respect to multiplication or survival in mice (Whittle et al. in press). In IL5 deficient mice, IgA titres against LPS in intestinal secretions from vector-only immunised mice were at least twofold lower than in normal mice. This difference was abolished by the immunisation with the IL5 secreting recombinant bacteria (Whittle et al. in press) supporting the suggestion that IL5 plays an enhancing role in IgA production in vivo. These data are in contrast to those obtained after intranasal infection of the respiratory tract with influenza virus where no differences in IgA producing B cells were found between IL5 deficient and normal mice (Kopf et al. 1996). The reason for these differences is not yet clear but it is worth noting that other studies have also supported a role for IL5 in these processes (Kinashi et al. 1986, Murray et al. 1987, Beagley et al. 1988). B-1 B lymphocyte development is IL-5 dependent B-1 B lymphocytes are considered to be the major source of IgM serum levels and are the first line of defence in natural immunity (Kocks & Rajewsky 1989, Kearney et al. 1992, Hardy & Hayakawa 1994). B-1 B cells possess IL5 receptors (Wetzel 1989) however early studies on B-1 B lymphocytes in IL5^-/- mice indicated that there was only a transient reduction of B-1 B cells at two weeks of age returning to normal levels in the adult deficient mice (Kopf et al. 1996). In contrast more recent studies (Bao et al. in preparation), show a 50% reduction of B-1 B cells (IgA^+/CD5lo and IgM^+/CD5lo) in the intestinal lamina propria and peritoneal cavity of IL-5^-/- adult mice, compared with controls. Functionally, the reduction in B-1 cells correlated with an approximate 50% reduction of anti-phosphatidyl choline (B-1 antigen) antibody levels in faecal pellets and the intestinal washings from both naive and S. typhimurium-challenged IL5^/- mice (Bao et al. in preparation). However, no significant differences of anti-OVA (B-2 antigen) antibody levels in both faecal pellets and intestinal washings were found in both groups of mice. These findings suggest that the development of B-1 B cells, but not B-2 B cells, are IL-5 dependent (Bao et al. in preparation) This is contrary to earlier findings in the IL5^-/- mice (Kopf et al. 1996) but is consistent with results obtained with transgenic mice overproducing IL5 (Tominaga et al. 1991) and in mice lacking the IL5 receptor (Yoshida et al. 1996). Conclusion IL5 deficient mice provide a convenient and informative model for studying the role of IL5 and eosinophils in vivo. Although basal levels of eosinophils are present in these animals, the absence of eosinophilia in the IL5^-/- mice provides the opportunity to test whether eosinophils have an important role in processes such as host defence against parasites, in diseases such as asthma and in a number of other biological situations where a local accumulation of eosinophils is observed. In parasite infections there is evidence for a role of IL5 and eosinophils in host defence against S. ratti, H. polygyrus as well as T. spiralis, but not against M. corti or T. canis. IL5 and eosinophils also play a major role in aeroallergen-induced lung damage and the development of airways hyper-responsivenes in a mouse model of asthma. Further support has also been obtained for a role of IL5 in the regulation of peritoneal B-1 cells and in the production of IgA responses. However, studies thus far suggest that IL5 and eosinophils play no obligatory role in xenograft rejection, in disease progression in an ulcerative colitis model or in uterine function and fertility. Continued detailed studies on the IL5 deficient mouse should enhance our understanding of IL5 and eosinophil function in vivo. Acknowledgements To our collaborators Bob Bao and Alan Husband; Carol Behm and Karen Ovington; Arne Mould; Sarah Robertson; Liljana Stevceva and Paul Pavli; as well as Bruce Vallence and Steven Collins for allowing us to refer to as yet unpublished data. To Elizabeth Milbourne and Carol Behm for infecting the mice with Mesocestoides corti used for analysis by electron microscopy. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97170a.jpg] |
| |||||||||

{kind=link}
{kind=link}