 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Pulmonary Biology of Anti-interleukin 5 Antibodies
RW Egan^+, D Athwahl, C-C Chou, RW Chapman, S Emtage, C-H Jenh,
Schering-Plough Research Institute, 2015 Galloping Hill Road, Kenilworth,
NJ 07033, USA and Celltech Therapeutics, 261 Bath Road, Slough, SL 14 FN,
U.K. Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97171
Sizes of Files:
Text: 18.3K
Graphics: Line drawings and photographs (jpg) - 112.1K
Tables (jpg) - 20.9K
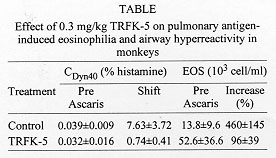
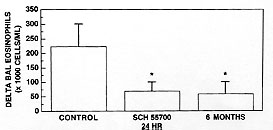
Interleukin 5 (IL-5) is a critical cytokine for the maturation of eosinophil precursors to eosinophils in the bone marrow and those eosinophils then accumulate in the lungs during asthma. We have studied anti IL-5 antibodies on allergic responses in mice, guinea pigs and monkeys and are extending this experiment into humans with a humanized antibody. In a monkey model of pulmonary inflammation and airway hyperreactivity, we found that the TRFK-5 antibody blocked both responses for three months following a single dose of 0.3 mg/kg, i.v. This antibody also blocked lung eosinophilia in mice by inhibiting release from the bone marrow. To facilitate multiple dosing and to reduce immunogenicity in humans, we prepared Sch 55700, a humanized antibody against IL-5. Sch 55700 was also active against lung eosinophilia in allergic monkeys and mice and against pulmonary eosinophilia and airway hyperresponsiveness in guinea pigs. Furthermore, as opposed to steroids, Sch 55700 did not cause immunosuppression in guinea pigs. Studies with this antibody in humans will be critical to establishing the therapeutic potential of IL-5 inhibition. Key words: eosinophils - interleukin-5 - pulmonary inflammation - antibodies - airway hyperresponsiveness Pulmonary Biology of Anti-interleukin 5 Antibodies There is strong but circumstantial evidence for a role of interleukin-5 (IL-5) in asthma, because IL-5 is an essential cytokine for the maturation of eosinophil precursors to eosinophils and it is those eosinophils that accumulate in the lungs during pulmonary inflammation (Gleich et al. 1995). Inhibiting the actions of IL-5 should, therefore, block the maturation of eosinophil precursors to eosinophils and remove at least one of the major causes of asthma. We have investigated the effects of anti-IL-5 antibodies on allergic responses in mice, in guinea pigs and in monkeys and will be extending this experimental paradigm into man using a humanized antibody against IL-5. Using that antibody, we will acquire a good deal of concrete information regarding the role of IL-5 in asthma. The concept behind using an antibody to inhibit the action of a cytokine is that it binds the ligand making it inaccessible to its receptor. It should, therefore, bind the cytokine with a higher affinity than the receptor itself and, in the case of the antibodies described herein, such is the case. TRFK-5 antibody was raised against purified murine IL-5 (Schumacher et al. 1988) and has an IgG1 isotype. We have also prepared a CDR grafted humanized antibody against IL-5 named Sch 55700. Both of them have KD values of about 10^-10 M and both of them neutralize IL-5 from a variety of species. In our monkey model of asthma, adult cynomolgus monkeys were tranquilized with ketamine, injected intravenously with the antibody of choice, anesthetized and intubated with an endotracheal tube through which we inserted a pediatric bronchoscope in order to conduct broncheoaveolar lavage with two 10 ml saline washes (Mauser et al. 1995). At the same time, we evaluated the responsiveness of the lungs to a dose response of aerosol histamine. These monkeys were naturally sensitive to aerosol Ascaris antigen, which was used as the allergic stimulus. Twenty-four hour later, bronchoalveolar lavage and airway hyperreactivity were evaluated again such that each monkey served as its own control. In a set of six monkeys, airway hyperreactivity and bronchoalveolar lavage eosinophils were measured in the absence of the antibody. Two weeks later, when the pulmonary parameters in this set of monkeys had returned to baseline, the experiment was repeated in the presence of the antibody, such that this set of monkeys was internally controlled. To quantitate airway hyperreactivity, we measured the histamine concentration required to cause a 40% decrease in dynamic compliance, the CDyn40, and defined a shift ratio as the CDyn40 before divided by the CDyn40 after Ascaris challenge. A shift greater than 1 indicates hyperreactivity while a shift of 1 indicates that the status of the lungs is the same before and after the allergic challenge. Using this experimental protocol, we examined the effects of 0.3 mg/kg of the TRFK-5 antibody on Ascaris induced pulmonary eosinophilia and airway hyperreactivity in monkeys (Table) (Mauser et al. 1995). In the control experiment, we treated the six animals with histamine to establish the pre-Ascaris CDyn40, which turned out to be 0.04% histamine. Subsequent to the allergic challenge, we measured the response to histamine again and established a shift ratio of 7.6 fold. In these monkeys, lavage eosinophil levels were 14 x 10^3/ml before the allergic challenge and 4-5 fold higher after the Ascaris challenge. Two weeks later when these animals had largely returned to normal, the same experiment was performed in the presence of 0.3 mg/kg TRFK-5 antibody. The pre-Ascaris CDyn40 was 0.03 %, virtually the same as previously. However, instead of the shift of 7-8 fold, the shift ratio was 0.74, indicating that the monkey's lungs were not hyperreactive. Baseline eosinophils were about 52,000/ml, but there was only about a 2-fold increase. Irrespective of whether the decrease in lung eosinophilia caused the reduced hyperreactivity, anti-IL-5 treatment inhibited the airway hyperreactivity and this physiology should translate into meaningful results in human asthma.
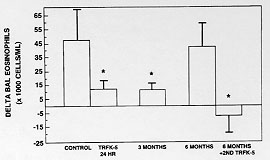
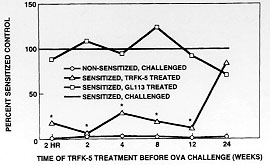
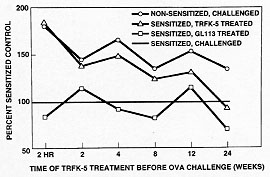
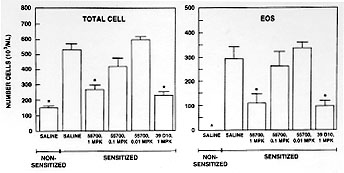
In order to determine whether this phenomenon was peculiar to the monkey and to extend the experimental paradigm into a species that is more experimentally tractable, we have studied the extended duration of action of the TRFK-5 antibody in the B6D mouse. Mice were sensitized to alum precipitated ovalbumin by a single 8 mg intraperitoneal injection. Five days later, they were boosted with the same dose to enhance the responsiveness. After 12 days, when IgE levels had increased about 7-fold, they were challenged with aerosol ovalbumin, causing an allergic response. Twenty-four hours after the challenge the mice were sacrificed and specimens of lavage fluid, lung tissue, blood and bone marrow were analyzed for total cells and eosinophils. Antibody was administered 2 hr prior to the ovalbumin challenge.
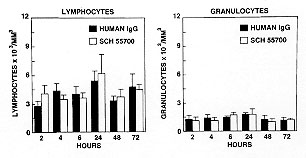
To conduct similar studies in the human, we would need a humanized antibody against IL-5 to facilitate multiple dosing and to reduce immunogenicity. We, therefore, performed CDR grafting based on the 39D10 antibody. The CDR's from 39D10 were grafted onto an otherwise human framework with an IgG4 isotype and a few framework residues in the region of the CDR's in order to achieve maximum binding affinity. This antibody, named Sch 55700, was first tested for its innate activity to bind human IL-5 compared to 39D10 (Egan et al. 1995). Using a BIAcore binding assay, the on rates for both 39D10 and Sch 55700 were virtually identical at 5 x 10^5M^-1sec^-1. The off rates between 3 and 4 x 10^-5sec^-1 gave KD values of 5 and 8 x 10^-11M, respectively. To evaluate the neutralizing capacity, we tested antibody based inhibition of IL-5 induced proliferation of the TF-1 human erythroleukemic cell line. Again, both antibodies were active in the range of 4 x 10^-11M. Hence, Sch 55700 is as potent as 39D10, and this binding affinity is stronger than the association between IL-5 and its high affinity receptor, which has a KD of about 25 x 10^-11M.
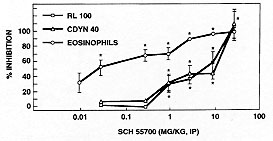
Sch 55700 was also examined in guinea pigs, because it was possible to study both pulmonary eosinophilia and airway hyperreactivity in that species. The experimental paradigm was very similar to the mouse. The guinea pigs were sensitized to intraperitoneal ovalbumin then challenged with aerosol ovalbumin, following treatment with the antibody or with a sham injection. We then examined lung tissue eosinophilia, lavage eosinophilia and reactivity of the guinea pigs airways to substance P. Fig. 6 shows the effects of intraperitoneal Sch 55700 on both eosinophil infiltration and airway hyperreactivity as indicated by either CDyn40 or specific lung resistance (RL100). The eosinophil infiltrate was inhibited by 0.03 mg/kg, while it took 1 mg/kg before the physiology was altered. Both the eosinophil infiltration and the airway hyperreactivity were blocked completely at high doses of Sch 55700.
Acknowledgment To Terri Ruhnke for preparing the manuscript. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97171f.jpg] [oc97171d.jpg] [oc97171e.jpg] [oc97171g.jpg] [oc97171a.jpg] [oc97171b.jpg] [oc97171c.jpg] [oc97171h.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}