
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 92, Num. s2, 1997, pp. 75-91
|
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92, Suppl. II, pp. 75-91
IL-5 and IL-5 Receptor in Asthma
ATC Kotsimbos, Q Hamid^+
Department of Medicine, Meakins-Christie Laboratories, McGill University,
3626 rue St Urbain, Montreal, Quebec, Canada H2X 2P2
^+Corresponding author. Fax: +514-398-7483. E-mail: Hamid@meakins.lan.mcgill.ca
Supported by MRC Canada and JT Costello Memorial Research fund.
Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97172
Sizes of Files:
Text: 88.7K
Graphics: Line drawings and photographs (jpg) - 98K
Eosinophils, along with mast cells are key cells involved in the innate
immune response against parasitic infection whereas the adaptive immune
response is largely dependent on lymphocytes. In chronic parasitic disease
and in chronic allergic disease, IL-5 is predominantly a T cell derived
cytokine which is particularly important for the terminal differentiation,
activation and survival of committed eosinophil precursors. The human IL-5
gene is located on chromosome 5 in a gene cluster that contains the
evolutionary related IL-4 family of cytokine genes. The human IL-5 receptor
complex is a heterodimer consisting of a unique alpha subunit (predominantly
expressed on eosinophils) and a beta subunit which is shared between the
receptors for IL-3 & GM-CSF (more widely expressed). The a subunit is
required for ligand-specific binding whereas association with the b subunit
results in increased binding affinity. The alternative splicing of the
alphaIL-5R gene which contains 14 exons can yield several alphaIL-5R
isoforms including a membrane-anchored isoform (alphaIL-5Rm) and a soluble
isoform (alphaIL-5Rs). Cytokines such as IL-5 produce specific and
non-specific cellular responses through specific cell membrane receptor
mediated activation of intracellular signal transduction pathways which, to
a large part, regulate gene expression. The major intracellular signal
transduction mechanism is activation of non-receptor associated tyrosine
kinases including JAK and MAP kinases which can then transduce signals via
a novel family of transcriptional factors named signal transducers and
activators of transcription (STATS). JAK2, STAT1 and STAT 5 appear to be
particularly important in IL-5 mediated eosinophil responses.
Asthma is characterized by episodic airways obstruction, increased
bronchial responsiveness, and airway inflammation. Several studies have
shown an association between the number of activated T cells and
eosinophils in the airways and abnormalities in FEV1, airway reactivity and
clinical severity in asthma. It has now been well documented that IL-5 is
highly expressed in the bronchial mucosa of atopic and intrinsic asthmatics
and that the increased IL-5 mRNA present in airway tissues is predominantly
T cell derived. Immunocytochemical staining of bronchial biopsy sections
has confirmed that IL-5 mRNA transcripts are translated into protein in
asthmatic subjects. Furthermore, the number of activated CD 4 + T cells and
IL-5 mRNA positive cells are increased in asthmatic airways following
antigen challenge and studies that have examined IL-5 expression in
asthmatic subjects before and after steroids have shown significantly
decreased expression following oral corticosteroid treatment in
steroid-sensitive asthma but not in steroid resistant and chronic severe
steroid dependent asthma. The link between T cell derived IL-5 and
eosinophil activation in asthmatic airways is further strengthened by the
demonstration that there is an increased number of alphaIL-5R mRNA positive
cells in the bronchial biopsies of atopic and non-atopic asthmatic subjects
and that the eosinophil is the predominant site of this increased
alphaIL-5R mRNA expression. We have also shown that the subset of activated
eosinophils that expressed mRNA for membrane bound a IL5r inversely
correlated with FEV1, whereas the subset of activated eosinophils that
expressed mRNA for soluble alphaIL5r directly correlated with FEV1. Hence,
not only does this data suggest that the presence of eosinophils expressing
alphaIL-5R mRNA contribute towards the pathogenesis of bronchial asthma,
but also that the eosinophil phenotype with respect to alphaIL-5R isoform
expression is of central importance. Finally, there are several animal, and
more recently in vitro lung explant, models of allergen induced
eosinophilia, late airway responses(LARS), and bronchial
hyperresponsiveness(BHR) - all of which support a link between IL-5 and
airway eosinophila and bronchial hyperresponsiveness. The most direct
demonstration of T cell involvement in LARS is the finding that these
physiological responses can be transferred by CD4+ but not CD8+ T cells in
rats. The importance of IL-5 in animal models of allergen induced bronchial
hyperresponsiveness has been further demonstrated by a number of studies
which have indicated that IL-5 administration is able to induce late phase
responses and BHR and that anti-IL-5 antibody can block allergen induced
late phase responses and BHR.
In summary, activated T lymphocytes, IL5 production and eosinophil
activation are particularly important in the asthmatic response. Human
studies in asthma and studies in allergic animal models have clearly
emphasised the unique role of IL-5 in linking T lymphocytes and adaptive
immunity, the eosinophil effector cell, and the asthma phenotype. The
central role of activated lymphocytes and eosinophils in asthma would argue
for the likely therapeutic success of strategies to block T cell and
eosinophil activation (eg steroids). Importantly, more targeted therapies
may avoid the complications associated with steroids. Such therapies could
target key T cell activation proteins and cytokines by various means
including blocking antibodies (eg anti-CD4, anti-CD40, anti-IL-5 etc),
antisense oligonucleotides to their specific mRNAs, and/or selective
inhibition of the promoter sites for these genes. Another option would be
to target key eosinophil activation mechanisms including the alphaIL5r. As
always, the risk to benefit ratio of such strategies await the results of
well conducted clinical trials.
Key words: interleukin 5 - asthma - eosinophils
Asthma and allergic inflammation
Asthma is characterized by episodic airway obstruction, increased bronchial
responsiveness to the inhalation of non-specific irritants and airway
inflammation (McFadden & Gilbert 1992). The link between abnormal airway
physiology and airway inflammation was initially suggested by the results
of post-mortem studies of asthmatic lungs which documented the presence of
an inflammatory infiltrate (Dunnill 1960). The use of fibreoptic
bronchoscopy has allowed bronchial biopsies and lavage fluid to be examined
in less severe asthmatics and in normal subjects. These studies have shown
that eosinophils and T lymphocytes in particular are increased in number
and activation status in asthmatic airways compared to controls (Jeffery et
al. 1989, Azzawi et al. 1990, Bradley et al.1991, Bentley et al. 1992,
Laitinen et al. 1993). Furthermore, several studies have correlated the
number of activated T cells and eosinophils with abnormalities in FEV1 and
airway reactivity (Walker et al. 1991) and with clinical severity in asthma
(Bousquet et al. 1990).
More recently, lung resection studies have also demonstrated an increase in
the number of eosinophils and T lymphocytes in both the large and small
airways of asthmatic subjects compared to non-asthmatic controls matched
for age, sex, smoking history, lung function and airway size (Hamid et al.
1996). These results extend the findings from previous studies using
endobronchial biopsies by showing that a similar but more severe
inflammatory process is present in the peripheral airways and in the airway
wall external to the smooth muscle layer-both of which are not routinely
biopsied during fibreoptic bronchoscopy. This data is consistent with the
evidence that suggests that the small airways are the major site of
obstruction in asthma (Macklem et al. 1970, Wiggs et al. 1992, Kuwano et
al. 1993). In addition, the extensive presence of inflammatory cells
throughout the airways makes it possible that these cells may be important
modulators of the function of other cells present in airway
tissues-including epithelial cells, fibroblasts and smooth muscle cells.
There is therefore considerable interest and research into the mechanisms
underlying the initiation and maintenance of the inflammatory respone in
asthma-which is likely to be a consequence of a complicated interaction
between various cells and mediators.
IL-5: AN IMPORTANT LINK BETWEEN T CELLS and EOSINOPHILS
IL-5 molecular biology
IL-5 gene, mRNA and protein
The human IL-5 gene is located on chromosome 5 in a gene cluster that
contains the IL-4 family of cytokine genes (Boulay & Paul 1992). It
contains 4 exons which encode a peptide of 124 amino acids (Azuma et al.
1986). IL-5 is a monomer which exists functionally as an antiparallel
homodimeric glycoprotein linked by 2 disulfide bonds and has a tertiary
crystalline structue consisting of 4 alpha helices (Milburn et al. 1993).
The exon structure, primary secondary and tertiary protein sequences, cell
membrane receptors and intracellular signal transduction pathways of IL-5
are similar with those from the IL-4 cytokine family, thereby suggesting
that they are evolutionary related cytokines (Milburn et al. 1993, Kosugi
et al. 1995). Therefore, it is not surprising that these cytokines also
share cellular sources and functional activities and are all important in
the co-ordinated immune defense against parasitic infection. However, IL-5
is unusual in that it is the most highly conserved member of this group. In
addition, the IL-5 glycoprotein is highly homologous between mammalian
species thereby suggesting that IL-5 function is of particular benefit to
the host organism.
IL-5 receptor gene, mRNA and protein
The human IL-5 receptor complex is a heterodimer consisting of a unique a
subunit (alphaIL-5R; MW 60Kd) and a b subunit (MW 130Kd) which is shared
between the receptors for IL-3 and GM-CSF (Lopez et al. 1991, Murata et al.
1992, Miyajima et al. 1993). In vitro, the expression of human alphaIL-5R
has been described to be present on eosinophils and basophils whereas the b
subunit is more widely expressed (Denburg et al. 1991, Migita et al.1991,
Miyajima et al. 1993). The a subunit is required for ligand-specific
binding whereas association with the b subunit results in increased binding
affinity (Takagi et al. 1995). The receptors for IL-5, IL-3 and GM-CSF
belong to the class I cytokine receptor family based on their structural
motifs (Bazan 1990, Boulay & Paul, 1992b). The membrane proximal region of
the extracellular domains of both the a and b subunits of the IL-5R have
common structural features which they share with the other members of the
haematopoietin cytokine receptor family. This homologous region is
characterized by a trp-ser-x-trp-ser motif and by several conserved short
sequence elements - the integrity of which is required for the interaction
with its ligand (Bazan 1990, Boulay & Paul 1992b).
The gene for human alphaIL-5R is located on chromosome 3 (Tuypens et al.
1992) and the gene for human bIL-5R is on chromosome 22 (Miyajima et al.
1993, Takai et al. 1994). The alternative splicing of the alphaIL-5R gene
which contains 14 exons can yield several alphaIL-5R isoforms including a
membrane-anchored isoform (alphaIL-5Rm) and a soluble isoform (alphaIL-5Rs)
(Tuypens et al. 1992, Tavernier et al. 1992). The membranous and soluble
alphaIL-5R isoform primarily differ in whether or not a transmembrane
binding domain is present. Although, alphaIL-5Rm and alphaIL-5Rs isoform
bind IL-5 with equally high affinity (Tavernier et al. 1992, Devos et al.
1993, Koike et al. 1994), alphaIL-5Rm interacts with the b subunit thereby
substantially increasing the affinity for IL-5 and allowing specific signal
transduction pathways to be activated (Koike et al. 1994) whereas
alphaIL-5Rs competes for IL-5 ligand with alphaIL-5Rm present on
eosinophils and therefore has antagonistic properties that may have a
regulatory role (Tavernier et al. 1991).
IL-5 receptor mediated intracellular signal transduction
Cytokines produce specific and non-specific cellular responses through
receptor mediated activation of intracellular signal transduction pathways
which, to a large part, regulate gene expression (Nicola et al. 1989,
Miyajima et al. 1992). IL-5R mediated signalling requires the cytoplasmic
domains of both subunits, is dependent on the proline rich areas proximal
to the transmembrane domains and involves the process of dimerization of
the a and b subunits (Sakamaki et al. 1992, Miyajima et al. 1992, Takaki et
al. 1993). The major intracellular signal transduction mechanism is
activation of non-receptor associated tyrosine kinases including MAP
kinases (Matsumoto et al. 1995, Pazdrak et al. 1995) and JAKS (Sakamaki et
al. 1992, Corneils et al. 1995) which can then transduce signals via a
novel family of transcriptional factors named signal transducers and
activators of transcription (STATS) (Ihle et al. 1995a,b). STAT proteins
exist in the cytoplasm as latent, transcriptionally inactive forms until in
response to extracellular signals, they become phosphorylated on tyrosine
residues, translocate to the nucleus, and bind to specific DNA elements.
JAK2, STAT1 and STAT 5 appear to be particularly important in IL-5 mediated
eosinophil responses (Mui et al. 1995, Van der Braggen et al. 1995).
Despite major advances in this area in recent years, the exact mechanisms
by which IL-5 dependent, cell-type specific signals are generated are still
to be elucidated.
IL-5, T cells and eosinophils
Eosinophils, along with mast cells are the key cells involved in the innate
immune response against parasitic infection. The adaptive immune response
however is largely dependent on lymphocytes. CD4 +ve T lymphocytes, in
particular, are crucial in antigen-driven inflammatory processes and are
therefore likely to have an important role in orchestrating specific
inflammatory responses. These cells are capable of recognizing foreign
antigen that has been processed by antigen presenting cells and can produce
pro-inflammatory cytokines in response to such activation which can
dramatically amplify the inflammatory response.
One of the major links between T cells and eosinophils is IL-5. In chronic
parasitic disease and in chronic allergic disease, IL-5 is predominantly a
T cell derived cytokine whose major site of action is the eosinophil (Hamid
et al. 1991, Mahanty et al. 1993, Ying et al. 1995). Although the
development of tissue eosinophilia is T cell dependent, non T cell derived
IL-5 may also play an important role as IL-5 mRNA can also be produced by
mast cells and eosinophils (Plaut et al. 1989, Brodie et al. 1992). IL-5,
IL-3, and GM-CSF are all capable of stimulating the development of
eosinophils from human bone marrow. However, only IL-5 was selective for
the eosinophil lineage (Clutterbuck et al. 1989). Transgenic mice which
constituitively express IL-5 have high level, life-long eosinophilia (Dent
et al. 1990) and the administration of anti-IL-5 neutralising antibody in
parasite infected mice totally blocks the production of eosinophilia
(Coffman et al. 1989, Egan et al. 1995).
IL-5 is particularly important for the terminal differentiation of
committed eosinophil precursors (Clutterbuck et al. 1989, Weller et al.
1992, Ogawa 1994). It activates mature eosinophils and prolongs their
survival in culture (Yamaguchi et al. 1988)-possibly via its ability to
delay apoptosis (Yamaguchi et al. 1991), as well as selectively enhancing
eosinophil degranulation, antibody-dependent cytotoxicity and adhesion to
vascular endothelium (Lopez et al. 1988, Fujisawa et al. 1990). IL-5
enhances the capacity of eosinophils to release LTC4 (Weller et al. 1992)
and also primes basophils, leading to increased histamine and LTC4
generation (Bischoff et al. 1990, Laviollette et al. 1995) and increases
synthesis of IgM, IgA, IgE by B cells costimulated with IL-4 (Pene et al.
1988, Purkerson & Isakson 1992). Although IL-5 on its own is minimally
chemoattractant for eosinophils, its ability to significantly enhance the
properties of stronger eosinophil chemoattractants such as Rantes and
Eotaxin is probably more important (Sanderson, 1992, Sedgwick et al. 1995,
Collins et al. 1995, Rothenberg et al. 1996).
IL-5, The TH2 cytokine profile and allergic inflammation
The production of IL-5 by T cells, like that of other TH2 cytokines, is
independently regulated (Kelso, 1995, Naora et al. 1995, Sewell et al.
1996). Although individual T cells have the capacity to produce a wide
range of cytokines, distinct T cell populations and cytokine profiles exist
in chronic allergic inflammatory diseases (Miyajima et al. 1992, Van
Straaten et al. 1994, Kay et al. 1995). There are a number of potential
explanations for this phenomenon. Firstly, as has already been mentioned,
the IL-4 family of cytokine genes is clustered on chromosome 5, have
related evolutionary pathways and are therefore likely to be regulated by
similar factors. Secondly, these factors are likely to co-exist in
particular microenvironments - particularly when inflammation is driven by
similar aetiological agents. And thirdly, these family of cytokines tend to
upregulate themselves and downregulate opposing groups of cytokines in an
attempt to generate a specific type of adaptive immune response (Modlin et
al. 1993, Jung et al. 1995). Indeed, cross-regulation of T helper cell
populations occurs and, in the extreme case, this may lead to the
development of relatively homogeneous Th1 and Th2 cell T cell population
phenotypes (Kelso 1995).
Th2 cell populations tend to produce IL-4, 5, 13 and are associated with
humoral immunity and allergy whereas TH1 cell populations tend to produce
IFN-g and IL-2 and are associated with cell mediated immunity (Modlin et
al. 1993). IL-4 and IL-13 are the cytokines that predominantly regulate B
cell production of IgE and IgE activation of mast cells, both of which have
an important role in the allergic immune response. The contribution of IL-4
and IL-5 to allergen induced eosinophil infiltration into the airway has
been suggested by experiments showing inhibition of airway eosinophilia in
mice with monoclonal antibodies directed against IL-4 and IL-5 (Moser et
al. 1992, Kung et al. 1995). These cytokines may act as chemotactic factors
for eosinophils, and also promote eosinophil-endothelial adhesion by
inducing expression of VCAM-1 on endothelial cells. VCAM -1 in turn may
bind to its receptor VLA-4 on the eosinophils leading to the migration of
eosinophils to sites of airway inflammation (Elices et al. 1990). However,
as has already been mentioned, one of the key roles of IL-5 however is to
regulate eosinophil activation, differentiation and survival. Thus,
although IL-5 also helps in the activation of B cells (Noelle et al. 1992),
its major role is to recruit and activate eosinophils which act in concert
with mast cells and IgE producing B cells in the immune response against
parasites and in pathophysiology of allergic disease. The relative
importance of these pathways is likely to vary according to the specific
`allergic' disease state. Nevertheless, the correlations between detectable
levels of IL-5 mRNA in the tissues and IL-5 protein in the serum,
eosinophilia development and disease pathology in a wide variety of
allergic diseases are striking (including parasite infections, asthma,
idiopathic eosinophilia, eosinophilic myalgia and Hodgkins lymphoma)
(Sanderson 1992b). Hence, activated T cells can potentially initiate and
propagate allergic inflammation in the airways and participate directly in
the events responsible for asthma exacerbation by profoundly influencing
both subsequent lymphocyte cell activation and the promotion of growth and
differentiation of specific effector leucocytes such as eosinophils.
ASTHMA and IL-5
HUMAN STUDIES: IL5 mRNA and protein
Atopic asthma - It has now been well documented that IL-5 is highly
expressed in the bronchial mucosa of atopic asthmatics and that the
increased IL-5 mRNA present in airway tissues is predominantly T cell
derived although fewer, but detectable, numbers of tryptase+ mast cells and
EG2+ eosinophils also expressed these transcripts (Hamid et al. 1991,
Robinson et al. 1992, Kay et al. 1995, Ying et al. 1995).
Immunocytochemical staining of bronchial biopsy sections has confirmed that
IL-5 mRNA transcripts are translated into protein in asthmatic subjects
(Fukuda et al. 1994).
Increased IL-5 mRNA expression has also been demonstrated in BAL T
lymphocytes (Robinson et al. 1992) and in peripheral blood CD4 T cells
(Corrigan et al. 1995). The reports of increased IL-5 protein levels in the
BAL fluid, serum and peripheral blood T cell supernatants of asthmatics
(Walker et al. 1992, 1991, Motojima et al. 1993), support the findings of
increased numbers of IL-5 mRNA positive cells in these biological fluids.
In addition, T cell lines that have been established from the BAL fluid and
peripheral blood of atopic asthmatics secrete increased levels of IL-5
compared to atopic and non-atopic controls, thus providing further evidence
that T cells in asthma have a propensity to make both IL-5 mRNA transcripts
and IL-5 translated product which can then activate eosinophils (Endo et
al. 1993, Okudaira et al. 1995, Till et al. 1995).
Numerous studies have shown that not only is there increased IL-5 mRNA and
protein present in asthma, but also that the increased IL-5 is associated
with increased eosinophil numbers and increased airways dysfunction (Hamid
et al. 1991, Robinson et al. 1993, Sur et al. 1995). It has been
demonstrated that the number of IL-5 mRNA-positive cells correlates with
the number of eosinophils infiltrating the bronchial mucosa of asthmatic
subjects and that IL-5 expression inversely correlates with pulmonary
function (Hamid et al. 1991). In addition, the increased expression of IL-5
mRNA in BAL has been directly correlated to asthma symptom severity and
inversely correlated to abnormal airway physiology (Robinson et al. 1993).
Finally, in the studies where increased IL-5 protein levels were
demonstrated in the BAL fluid, serum and peripheral blood T cell
supernatants (Walker et al. 1992, 1991, Motojima et al. 1993) of asthmatic
subjects, the IL-5 protein levels detected correlated with the numbers of
eosinophils present in these fluids (Walker et al. 1992).
Activated eosinophils have the capacity to produce effector molecules that
could participate in the pathogenesis of asthma. The demonstration of
eosinophil major basic protein and eosinophil derived neurotoxin indicating
degranulation at sites of injury are an important part of the evidence that
eosinophils are producing tissue damage in the asthmatic lung (Sur et al.
1995). Hence, the current evidence suggests that the local production of
IL-5 in asthmatic airways may play an important role in the priming of
eosinophils for subsequent activation, and in enhancing their survival at
sites of allergic inflammation (Lopez et al. 1988, Yamaguchi et al. 1988),
all of which is likely to be important in asthma.
Intrinsic asthma - Unlike extrinsic asthma, intrinsic asthma usually
starts in adulthood, is perennial and is not atopy associated.
Nevertheless, an analysis of the inflammatory cell populations present in
both BAL fluid and bronchial biopsies from intrinsic asthmatics shows an
increase in the number of activated T lymphocytes and eosinophils (Bentley
et al. 1992). Indeed, there a large similarities in the inflammatory cells
that are present in asthma of diverse aetiology-extrinsic, intrinsic, and
occupational (Bentley et al. 1994). Moreover, several studies have now
demonstrated increased levels of IL-5 mRNA and protein in the tissue and
BAL fluid of intrinsic asthmatics, thereby supporting the role of this
cytokine in both intrinsic as well as atopic asthma (Marini et al. 1992,
Bentley et al. 1993, Walker et al. 1994, Humbert et al. 1996). These
findings support a common T cell mediated basis for airway inflammation in
both forms of asthma.
The difference between intrinsic and atopic asthma is the lack of
demonstrable specific IgE to an antigen in individuals with intrinsic
disease. Although total serum IgE levels have been noted to be increased in
the serum of patients with intrinsic asthma (Burrows et al. 1989) this is
not a uniform finding with high positive predictive value (Butcher et al.
1980, Klink et al. 1990). There is evidence that IL5 but not IL-4 is
increased in the BAL fluid from intrinsic asthmatics (Walker et al. 1994),
thus supporting the hypothesis that IL-5 and eosinophilia are key features
in both forms of asthma and that differences in the type of cytokine
synthesis may undermine the differences in immune pathology that exist
between intrinsic and atopic asthma. However, more recent studies examining
the expression of high-affinity IgE receptor (Humbert et al. 1996a), and
IL-5 and IL-4 mRNA and protein expression in bronchial biopsies from
patients with atopic and non-atopic asthma found no difference between
atopic and intrinsic asthmatics (Humbert et al. 1996b). Thus, any
differences in immune pathology that may exist between intrinsic and atopic
asthma may be more subtle than initially expected.
Other allergic pulmonary and non-pulmonary allergic diseases - There
is a strong association between IL-5 and eosinophilia in a number of
allergic human diseases, including parasitic infections (Coffman et al.
1989, Limaye et al. 1993, Hagan et al. 1996), atopic dermatitis (Frew & Kay
1988, Hamid et al. 1994), eosinopilic myocarditis (Desreumaux et al. 1993),
hypereosinopilic syndrome (Schrezenmeier et al. 1993, Satoh et al. 1994)
eosinophilic gastroenteritis (Quan et al. 1993, Dubucquoi et al. 1995),
allergic rhinitis (Durham et al. 1992), chronic eosinophilic pneumonia
(Kita et al. 1996) and other eosinophilic lung diseases (Walker et
al. 1994) (Fig. 1).
Figure 1: IL-5 in
asthma and other pulmonary diseases.
The percentage of IL-5 mRNA +ve cells in the BAL fluid of patients with
asthma, sarcoidosis, and tuberculosis. NC: normal controls; NA: non-active
disease; A: active disease; NS: non-symtomatic disease; S: symptomatic
disease.
Although there is a strong link between IL-5 and eosinophilia in general,
the link between pulmonary eosinophilia and clinical asthma is less direct.
Non-eosinophilic inflammatory lung conditions such as tuberculosis and
sarcoidosis are not associated with increased IL-5 and eosinophils (Taha et
al. 1996, Minshall et al. 1996). However, although eosinophilia is a common
feature of asthma many eosinophilic lung diseases are not associated with
clinical asthma. This argues that factors other than the presence of
eosinophils are also important in the development of clinical asthma. These
factors may relate to the level of eosinophil activation, to non-eosinophil
dependent parameters that are nevertheless associated with allergic
inflammation, or to baseline levels of bronchial hyperresponsivenes. It is
therefore likely that the asthma phenotype is most likely to occur when all
the relevent factors- including IL5 and eosinophilia, occur together in an
individual predisposed to bronchial hyperresponsiveness.
Figure 2: IL-5 mRNA
following allergen challenge
humans. The number of IL-5 mRNA +ve cells/mm basement membrane in atopic
asthmatic subjects following specific allergen challenge.
Antigen challenge - The number of activated CD 4 + T cells and IL-5
mRNA positive cells are increased in asthmatic airways following antigen
challenge (Robinson et al. 1993, Bentley et al. 1993) (Fig. 2).
Furthermore, CD 4 + ve T cells have been generally implicated as the major
IL-5 mRNA positive cell present following antigen challenge in atopic
asthmatics (Bentley et al. 1993, Robinson et al. 1993), although some
investigators have reported that the eosinophil is also a source of IL-5 in
this setting (Broide et al. 1992). Studies examining BAL samples
18-48 hr after allergen challenge have also shown increased expression of
IL-5 (Krishnaswamy et al. 1993, Ohnishi et al. 1993). In addition, IL-5 was
a major cytokine product of T cells from patients with mite associated
bronchial asthma when they were stimulated with Dermatophagoides farinae
(Kamei et al. 1993).
The increased IL-5 expression that follows allergen challenge has been
demonstrated to inversely correlate with pulmonary function (Bentley et al.
1993) and this adds to the evidence that IL-5 expression and eosinophilia
are relevently increased following exposure to antigen in sensitised
individuals. Indeed, in the study by Ohnishi et al. (1993)
a segmental antigen lung challenge model was used to show that IL-5 was
the most important constituent increasing eosinophil survival and that IL-5
correlated with eosinophil recruitment, degranulation and lung injury
following inhalation of antigen. These results are in agreement with
several other studies that have indicated that increases in the levels of
eosinophils and their cationic proteins in the BAL fluid following allergen
challenge correlates with the magnitude of the late phase response
(Pradalier 1993).
Furthermore, it is well recognized that there is an association between
allergic rhinitis and allergic asthma, and hence studies using models of
allergen-induced allergic rhinits are therefore relevent to allergic
asthma. The results obtained from such models by and large support the
above findings. For instance, it has been shown that T cells are the
principal source of IL-5 transcripts in the nasal mucosa following allergen
induce late-phase nasal responses (Ying et al.1993). Similar findings have
also been reported in models of allergen induced cutaneous late phase
reactions (Kay et al. 1991).
Steroid treatment - The use of anti-inflammatory corticosteroids are
the cornerstone of current asthma therapy. They have been shown to be
extremely effective clinically. Studies that have examined IL-5 expression
in BAL and peripheral blood of asthmatic subjects before and after steroids
have shown that the number of IL-5 mRNA positive cells is significantly
decreased following oral corticosteroid treatment in steroid-sensitive
asthma (Robinson et al. 1993, Corrigan et al. 1995). In contrast, steroid
resistant asthma and chronic severe steroid dependent asthma are associated
with persistently elevated IL-5 mRNA levels (Leung et al. 1995) and serum
IL-5 levels (Alexander et al. 1994), respectively . The decreases in the
expression of IL-5 that followed corticosteroid therapy have been
associated with decreased eosinophil numbers - especially in the peripheral
blood (Corrigan et al. 1995), but increased numbers of IFN-g positive cells
in the bronchial mucosa and BAL fluid of asthmatic subjects (Robinson et
al. 1993, Leung et al. 1995, Bentley et al. 1996). These findings support
the direct link between IL-5 and eosinophils and the inverse relationship
between Th1 and Th2 type T cells in asthma (Fig. 3). Hence, corticosteroid
treatment in asthma may act by modulation of cytokine expression with
consequent inhibition of the local bronchial inflammatory infiltrate and
tissue eosinophilia.
Figure 3: prednisone
therapy and IL-5 mRNA expression
in steroid-sensitive and steroid-resistant asthmatics. The number of IL-5
mRNA +ve cells/mm basement membrane in the bronchial mucosa before and
after prednisolone therapy in steroid-sensitive and steroid-resistant
asthmatics.
IL5 RECEPTOR (Membrane bound and soluble isoforms)
The link between T cell derived IL-5 and eosinophil activation in asthmatic
airways has now been supported by the demonstration that there is an
increased number of alphaIL-5R mRNA positive cells in the bronchial
biopsies of atopic and non-atopic asthmatic subjects compared with atopic
and non-atopic controls (Fig. 4a, b) and that the eosinophil is the
predominant site of this increased alphaIL-5R mRNA expression (Yasruel et
al. 1997). We have shown that 93% of the alphaIL-5R mRNA positive cells
within the bronchial mucosa of asthmatics were also EG2 positive thereby
suggesting that IL-5 may play an important local role in stimulating
eosinophils via the specific a-subunit of its receptor. These results
support previous work that has suggested that the lineage specificity of
IL-5 is mainly due to the restricted expression of the a subunit of IL5R
(Takagi et al. 1995) and demonstrates that the expression of alphaIL-5R in
vivo can be much more cell-restricted than that seen in vitro (Lopez et al.
1991).
Figure 4 a/b :
membrane and soluble IL-5R mRNA
expression in asthma. The number of membrane and soluble IL-5 receptor mRNA
+ve cells/mm basement membrane in the bronchial mucosa in asthma. mIL-5r:
membrane-bound IL-5r isoform; sIL-5r: soluble IL-5r isoform; NC: normal
controls; AC: atopic controls: AA: atopic asthmatics; IA: intrinsic
asthmatics.
We have also shown that the subset of activated eosinophils that expressed
mRNA for membrane bound IL5r inversely correlated with FEV1, whereas the
subset of activated eosinophils that expressed mRNA for soluble IL5r
directly correlated with FEV1. Hence, not only does this data suggest that
the presence of eosinophils expressing IL-5R mRNA contribute towards the
pathogenesis of bronchial asthma, but also that the eosinophil phenotype
with respect to alphaIL-5R isoform expression is of central importance.
Factors that may modulate the activation phenotype of the airway
eosinophils in asthma remain to be clearly determined. The apparent
contradiction between an EG2+ eosinophil (EG2 being a marker of activated
eosinophils) also expressing alphaIL-5Rs mRNA and hence representing a
downregulated cell can be explained by assuming that EG2 and the alphaIL5R
are associated with different levels of eosinophil activation. Indeed,
there is considerable controversy as to the validity of EG2 as a marker of
eosinophil activation (Moqbel et al. 1992). Moreover, there is no direct
way with which to grade the various potential activation of eosinophils. If
EG2 represented a relatively low grade eosinophil activation marker then it
is possible that when eosinophils cross the endothelial barrier they all
become EG2 + as a result of influences from the local tissue environment .
On the other hand, alphaIL-5R activation status might represent a higher
level of cell activation such that alphaIL-5Rm positive cells represented a
highly activated subset of EG2+ eosinophils and alphaIL-5Rs positive cells
a subset of EG2+ eosinophils that are minimally activated. Hence, although
previously published correlations between EG2+ eosinophils and FEV1 were
only modest (Hamid et al. 1991), such considerations may explain the strong
inverse correlation between alphaIL-5Rm mRNA positive cells and FEV1 as
well as the strong direct correlation between alphaIL-5Rs mRNA positive
cells and FEV1 that was reported.
The central question as to what controls the transcriptional regulation of
alphaIL-5R also remains to be determined. Transforming growth factor B1 has
already been shown to downregulate alphaIL-5R mRNA expression (Zanders
1994), however the effect of single cytokines or combinations of cytokines
on the production of different mRNA splice variants of alphaIL-5R is yet to
be elucidated. Nevertheless, the increased number of alphaIL-5R mRNA
positive eosinophils in the bronchial tissue of asthmatic patients and the
differential expression of alphaIL-5R mRNA isoforms in atopic and
non-atopic asthma support the central roles of IL-5 and eosinophils in the
pathobiology of asthma.
IL5 associated signal TX and gene activation
It has been shown that cloned human naive CD4 T cells develop into IL-4 and
IL-5 producing effector cells as a default pathway (Yang et al. 1995). It
could therefore be hypothesised that inert antigenic stimulation of the
immune system without concomitant stimulation of cell mediated immune
pathways would favour the development of allergic responses. However, how
exactly allergen induced activation of TCR and co-stimulatory molecules
translates to IL-4 and IL-5 gene activation is not clear. It could also be
hypothesised that it would be in the interests of a well coordinated
amplification cascade of inflammation to link IL-4 and IL-5 gene activation
in cytokine producing cells with IL-4 and IL-5 receptor gene activation in
target cells. Although we have already quoted some evidence that suggests
that this occurs, the exact mechanisms are unclear.
The functions of the alpha subunits of IL-5R and IL-4R have been examined
by co-transfecting human cDNAs for these subunits into human cell lines,
and it is clear that intracellular signalling is very different in both
cases (Chen et al. 1994). How IL-4 mediated intracellular signals interact
with the IL-5 gene promoter and whether IL-4 and IL-5 mediated signal
transduction can also increase the expression of IL-4 and IL-5 cytokines
from source cells and IL-4 and IL-5 receptors in target cells are important
issues still to be elucidated. Furthermore, how IL-5 mediated signals
translates to an activated eosinophil phenotype is also unclear at present,
although recent work has implicated specific GATA binding proteins (Zon et
al. 1993). Finally, the response of these cytokine and cytokine receptor
genes to therapeutic agents is another important area requiring further
study.
ANIMAL STUDIES
IL-5 EXPRESSION: sensitised and antigen challenged animals
There are several animal models of allergen induced eosinophilia, late
airway responses, and bronchial hyperresponsiveness. These include guinea
pigs (Corry et al. 1996), Brown norway (BN) rats (Renzi et al. 1991a,b,
1993, Olivenstein et al. 1993) and mice (Nakajima et al. 1992, Iwamoto et
al. 1992). In all these models there is evidence to support a link between
IL-5 and airway eosinophila and bronchial hyperresponsive-ness. In the BN
rat we have also shown that CD4+ve T cells and Th-2 cytokines, IL-5 in
particular, are involved in allergen induced late airway responses (LAR)
(Fig. 5) (Al Assad et al. 1995, Renzi et al. 1996).
Figure 5: IL-5 mRNA
expression following allergen
challenge in BN rats. The number of IL-5 mRNA +ve cells/mm basement
membrane in the bronchial mucosa of Brown Norway rats and Sprague-Dawley
rats following ovalbumin allergen challenge.
We have shown that the airways of OVA sensitized BN rats are infiltrated
predominantly by IL5 and IL4 mRNA +ve cells after antigen challenge (Renzi
et al. 1996). Eum et al. have demonstrated that eosinophil recruitment into
the respiratory epithelium following antigenic challenge is associated with
IL-5-dependent bronchial hyperresponsiveness (Eum et al. 1995). Recent work
has shown that IL-5 deficiency abolishes eosinophilia, airways
hyperreactivity and lung damage in a mouse asthma model and that
reconstitution of IL-5 production using recombinant vaccinia virus that
expressed IL-5 restored aeroallergen induced eosinophilia and airways
dysfunction (Foster et al. 1996). IL-5 transgenic mice show marked
eosinophilia and increased reactivity to acetylcholine only after antigen
challenge. This suggests that eosinopil activation rather than just large
numbers of eosinophils is crucial to the development of BHR (Iwamoto et al.
1995). Interestingly, genetic linkage analysis has linked bronchial
hyperesponsiveness in the mouse to murine chromosome 6 - the chromosomal
region containing the gene for IL-5 (Ewart et al. 1996).
Using an in vitro lung explant model it has also been shown that airways of
OVA sensitized BN rats are infiltrated predominantly by MBP + ve, IL5 and
IL4 mRNA +ve cells after ex-vivo antigen challenge (Fig. 6) (Minshall et
al. 1996). The demonstration of increased MBP and IL-5 mRNA expression in
sensitized lung explants after allergen challenge, suggests that local
factors are likely to be very important in the initiation and development
of airway eosinophil infiltration.
Figure 6: MBP and
IL-5 mRNA expression in sensitized BN
rat lung explants. The number of MBP and IL-5 mRNA +ve cells/mm basement
membrane in sensitised Brown Norway rat lung explant tissue before and
after ovalbumin allergen challenge. LAR: late phase airway response; BC:
before challenge; AC: after challenge.
The most direct demonstration of T cell involvement in LARs is the finding
that these physiological responses can be transferred by CD4+ but not CD8+
T cells in rats (Watanabe et al. 1995a,b). To investigate the role of T
cell cytokines in these responses the expression of mRNA for Th2 (IL-4 and
IL-5) and Th1(IL -2 and INF-g) type cytokines in BN rats that were
administered aerosolized OVA challenge following the adoptive transfer of
either antigen-primed W3/25(CD4)^+ or OX8(CD8)^+ T cells was examined (Fig.
7) (Watanabe et al. 1996). Our results showed that recipients of OVA-primed
CD4^+ T cells had an increase in the fraction of BAL cells expressing mRNA
for IL-4 and IL-5 compared to BSA-primed CD4^+ or OVA-primed CD8^+ cells.
Recipients of CD8^+ T cells had an increase in INF-g mRNA expression after
OVA challenge compared to recipients of BSA primed CD8^+ or OVA primed
CD4^+ T cells. Hence, T cell dependent allergen induced late responses are
associated with the expression of mRNA for IL-4 and IL-5, indicating Th2
cell activation. Furthermore, the increased expression of INF-g in allergen
challenge recipients of antigen-primed CD8^+ T cells suggests that CD8^+ T
cells may be important in modulating allergic responses, thus supporting
the conclusions from previous work in this area (Al Assad et al. 1995).
Figure 7: IL-5
mRNA-positive cells in BAL following
adoptive transfer of T cells. The number of IL-5 mRNA +ve cells/1000 cells
in the BAL fluid of OVA-challenged BN rats that were recipients of
OVA-primed T cells (CD4+ and CD8+groups).
IL-5 and anti-IL-5 administration
The importance of IL-5 in animal models of allergen induced bronchial
hyperresponsiveness has been further demonstrated by a number of studies
which have indicated that IL-5 administration is able to induce late phase
responses and BHR and that anti-IL-5 antibody can block allergen induced
late phase responses and BHR. IL-5 administration has been shown to
increases mucosal exudation, enhance eosinophil recruitment into the lungs
and to increases airways responsiveness in models of allergen induced BHR
in the guinea pig (Gulbenkian et al. 1992, van Oosterhout et al. 1993a), in
mice (van Oosterhout et al. 1993b), and in the Brown Norway rat (Renzi et
al. 1996). Moreover, anti-IL-5 administration inhibits eosinophil
recruitment and airways hyperresponsiveness in guinea pig models of
allergic pulmonary inflammation and allergen induced BHR (Gulbenkian et al.
1992, van Oosterhout et al. 1993a, Das et al. 1995). Similar findings have
also been demonstrated in the the mouse (Nagai et al. 1993, Kung et al.
1995) and in the monkey (Mauser et al. 1995).
IL-5 signal transduction modulation
Many animal studies have now demonstrated that eosinophilia is a uniquely
specific phenomenon regulated by IL-5 which of course suggest that IL-5
gene expression is under specific control. This control has been
investigated by examining the roles of various transcription factors in
animal T cell lines (Lee et al. 1994, Karlen et al. 1996). These studies
have implicated AP-1, NF-AT like factors and GATA binding proteins although
it is still not clear how all these molecules are related in controlling
IL-5 promoter activity (Lee et al. 1994, Yamagata et al. 1995, Karlen et
al. 1996).
THERAPEUTIC IMPLICATIONS
Asthma is a complex disorder involving a specific inflammatory response in
the airways that is largely co-ordinated by activated T cells and involves
various other inflammatory effector cells especially eosinophils but also B
cells and mast cells, as well as functional and structural changes in the
resident airway tissue cells. The central role of activated lymphocytes and
eosinophils in asthma would argue for the likely therapeutic success of
strategies to block T cell and eosinophil activation. It is likely that at
least some of the success of steroids in suppressing asthmatic inflammation
is due to their ability to suppress T lymphocyte and eosinophil activation.
However, steroids have a wide range of effects on many other inflammatory
and non-inflammatory cells. Although this may be the basis of the
therapeutic usefulness of steroids, it is also the basis of the significant
side effects associated with their long term use. Importantly, more
targeted therapies may avoid the complications associated with steroids.
Such therapies could target key T cell activation proteins and cytokines by
various means including blocking antibodies (eg anti-CD4, anti-CD40,
anti-IL-5 etc), antisense oligonucleotides to their specific mRNAs, and/or
selective inhibition of the promoter sites for these genes. Another option
would be to target key eosinophil activation mechanisms including the
alphaIL5r. As always, the risk to benefit ratio of such strategies await
the results of well conducted clinical trials.
The groundwork for such strategies is currently being laid. Anti-IL5
antibodies have been shown to inhibit pulmonary eosinophilia, tissue damage
and hypereactivity in allergic animal models (Egan et al. 1995, Hagan et
al. 1996). It has also been demonstrated that FK506 could suppress IL-5
production and gene expression in a dose dependent manner-(Okudaira et al.
1995). In addition, the data of Yasruel et al. (1996) linking soluble
alphaIL-5r expression with improved FEV1 levels and studies examining the
therapeutic potential of soluble alphaIL-5r suggest that they may offer
particular promise (Devos et al. 1995). Indeed, the possibility of the
antagonistic properties of the alphaIL-5Rs being used as a therapeutic
option in eosinophil mediated disease states such as asthma and allergic
rhinitis is already being investigated (Zanders 1994, Devos et al. 1995),
although the response of alphaIL-5R expression to antigen challenge and
steroid treatment are still important areas of further study.
Interestingly, the potential biological modulatory role of alphaIL-5Rs also
has important implications for the development of IL-5R antagonists as
these antagonists may not differentiate between binding to and inhibiting
alphaIL-5Rm, and binding to and inhibiting alphaIL-5Rs (Devos et al. 1994,
1995). Furthermore, in a murine model of allergic responses, it has been
shown that soluble alphaIL-5r suppressed antigen induced BAL eosinophilia
with little effect on airway hyperreactivity reminding us again of the
complexities involved in all these responses (Yamaguchi et al. 1994).
CONCLUSIONS
In summary, activated T lymphocytes, the production of IL-5 and eosinophil
activation are particularly important in the asthmatic response. Human
studies in asthma and studies in allergic animal models have clearly
emphasised the unique role of IL-5 in linking adaptive immunity and T
lymphocytes with the eosinophil effector cell. However, how this link
between IL-5 producing T cells and IL-5 target eosinophils is initiated,
propagated and attenuated is still an area that requires further research.
In addition, what the exact activation characteristics of the T lymphocytes
in asthma are (Vb restriction of their TCR in response to specific
antigens) and what the relative effects of T cells and Th2 cytokines are on
all effector inflammatory cells and on structural cells (epithelium,
fibroblasts and smooth muscle cells) of asthmatic airways are also
important issues that need to be resolved. It is hoped that answers to
these questions in the near future will provide us with an increased
understanding of asthma pathogenesis, and ultimately lead to novel, highly
targeted and effective therapeutic strategies for asthma management.
REFERENCES
Al Assad AS, Yang JP, Hamid Q, Yasruel Z, Renzi PM 1995.
Semi-quantitative analysis of cytokine mRNA in rat lungs after challenge.
Am J Resp Crit Care Med 151.4: (A392).
Alexander AG, Barkans J, Moqbel R, Barnes NC, Kay AB, Corrigan CJ 1994.
Serum interleukin 5 concentrations in atopic and non-atopic patients with
glucocorticoid-dependent chronic severe asthma. Thorax 49:
1231-1233.
Azuma C, Tanabe T, Konishi M, Kinashi T, Noma T, Matsuda F, Yaoita Y,
Takatsu K, Hammarstrom L, Smith CI 1986. Cloning of cDNA for human T-cell
replacing factor (interleukin-5) and comparison with the murine homologue.
Nucleic Acids Research 14: 9149-9158.
Azzawi M, Bradley B, Jeffery PK, Frew AJ, Wardlaw AJ, Assoufi B,
Collins JV, Durham SR, Knowles GK, Kay AB 1990. Identification of
activated T lymphocytes and eosinophils in bronchial biopsies in stable
atopic asthma. Am Rev Respir Dis 142: 1407-1413.
Bentley AM, Menz G, Storz C, Robinson DS, Bradley B, Jeffery PK, Durham
SR, Kay AB 1992. Identification of T lymphocytes, macrophages and
activated eosinophils in the bronchial mucosa in intrinsic asthma:
relationship to symptoms and bronchial responsiveness. Am Rev Respir
Dis 146: 500-506.
Bentley AM, Hamid Q, Robinson DS, Schotman E, Meng Q, Assoufi B, Kay
AB, Durham SR 1996. Prednisolone treatment in asthma. Reduction in the
numbers of eosinophils, T cells, tryptase-only positive mast cells, and
modulation of IL-4, IL-5, and interferon-gamma cytokine gene expression
within the bronchial mucosa. Am J Resp Crit Care Med 153:
551-556.
Bentley AM, Durham SR, Kay AB 1994. Comparison of the immunopathology
of extrinsic, intrinsic and occupational asthma. J Investig Allergy &
Clin Immunol 4: 222-232.
Bentley AM, Meng Q, Robinson DS, Hamid Q, Kay AB, Durham SR 1993.
Increases in activated T lymphocytes, eosinophils, and cytokine mRNA
expression for Interleukin-5 and granulocyte/macrophage colony-stimulating
factor in bronchial biopsies after allergen challenge in atopic
asthmatics. Am J Resp Cell Mol Biol 8: 35-42.
Bischoff SC, Brunner T, De Weck AL, Dahinden CA 1990. Interleukin 5
modifies histamine release and leukotriene generation by human basophils
in response to diverse agonists. J Exp Med 172: 1577-1582.
Bradley BL, Azzawi M, Jacobson M, Assoufi B, Collins JV, Irani AMA,
Schwartz LB, Durham SR, Jeffery PK, Kay AB 1991. Eosinophils,
Tlymphocytes, mast cells, neutrophils, and macrophages in bronchial biopsy
specimens from atopic asthma: comparison with biopsy specimens from atopic
subjects without asthma and normal control subjecs and relationship to
bronchial hyperresponsiveness. J Allergy Clin Immunol 88:
661674.
Bazan JF 1990. Structural design and molecular evolution of a cytokine
receptor superfamily. Proc Natl Acad Sci USA 87: 6934-6938.
Boulay JL, Paul WE 1992. The interleukin-4-related lymphokines and
their binding to hematopoietin receptors. J Biol Chem 267:
20525-20528.
Boulay JL, Paul WE 1992. The interleukin-4 family of lymphokines.
Curr Opin Immunol 4: 294-298.
Bousquet J, Chanez P, Lacoste JY, Barneon G, Ghavanian N, Enander I,
Venge P, Ahlstedt S, Simony-Lafontaine J, Godard P 1990. Eosinophilic
inflammation in asthma. New Engl J Med 323: 1033-1039.
Broide DH, Paine MM, Firestein GS 1992. Eosinophils express interleukin
5 and granulocyte macrophage-colony-stimulating factor mRNA at sites of
allergic inflammation in asthmatics. J Clin Invest 90:
1414-1424.
Burrows B, Martinez FD, Halonen M, Barbee RA, Cline MG 1989.
Association of asthma with serum IgE levels and skin-test reactivity to
allergens. New Engl J Med 320: 271-277.
Butcher BT, O'Neil CE, Reed MA, Salvaggio JE 1980. Radioallergosorbent
testing of toluene diisocyanate-reactive individuals using p-tolyl
isocyanate antigen. J Allergy & Clin Immunol 66: 213-216.
Chen JX, Watanabe S, Muto A, Miyajima A, Yokota T, Arai K 1994.
Activation of early response genes and cell proliferation by human
interleukin-3, granulocyte-macrophage colony-stimulating factor, and
interleukin-5 receptors: comparison with human interleukin-4 receptor
signaling. J Allergy & Clin Immunol 94: 605-611
Clutterbuck EJ, Hirst EM, Sanderson CJ 1989. Human interleukin-5 (IL-5)
regulates the production of eosinophils in human bone marrow cultures:
comparison and interaction with IL-1, IL-3, IL-6, and GMCSF. Blood
73: 1504-1512.
Coffman RL, Seymour BW, Hudak S, Jackson J, Rennick D 1989. Antibody to
interleukin-5 inhibits helminth-induced eosinophilia in mice.
Science 245: 308-310.
Collins PD, Marleau S, Griffiths-Johnson DA, Jose PJ, Williams TJ 1995.
Cooperation between interleukin-5 and the chemokine eotaxin to induce
eosinophil accumulation in vivo. J Exp Med 182:
1169-1174.
Cornelis S, Fache I, Van der Heyden J, Guisez Y, Tavernier J, Devos R,
Fiers W, Plaetinck G 1995. Characterization of critical residues in the
cytoplasmic domain of the human interleukin-5 receptor alpha chain
required for growth signal transduction. Eur J Immunol 25:
1857-1864.
Corrigan, CJ, Hamid Q, North J, Barkans J, Moqbel R, Durham S,
Gemou-Engesaeth V, Kay AB 1995. Peripheral blood CD4 but not CD8
T-lymphocyte in patients with exacerbation of asthma transcribe and
translate messenger RNA encoding cytokines which prolong eosinophil
survival in the context of a Th-2 type pattern: Effect of glucocorticoid
therapy. Am J Respir Cell Mol Biol 12: 567-578.
Corry DB, Folkesson HG, Warnock ML, Erle DJ, Matthay MA, Wiener-Kronish
JP, Locksley RM 1996. Interleukin 4, but not interleukin 5 or eosinophils,
is required in a murine model of acute airway hyperreactivity. J Exp
Med 183: 109-117.
Das AM, Williams TJ, Lobb R, Nourshargh S 1995. Lung eosinophilia is
dependent on IL-5 and the adhesion molecules CD18 and VLA-4, in a
guinea-pig model. Immunology 84: 41-46.
Denburg JA, Silver JE, Abrams JS 1991.Interleukin-5 is a human
basophilopoietin: induction of histamine content and basophilic
differentiation of HL-60 cells and of peripheral blood basophil-eosinophil
progenitors. Blood 77: 1462-1468.
Dent LA, Strath M, Mellor AL, Sanderson CJ 1990. Eosinophilia in
transgenic mice expressing interleukin 5. J Exp Med 172: 1425-1431.
Desreumaux P, Janin A, Dubucquoi S, Copin MC, Torpier G, Capron A,
Capron M, Prin L 1993. Synthesis of interleukin-5 by activated eosinophils
in patients with eosinophilic heart diseases. Blood 82:
1553-1560.
Devos R, Plaetinck G, Cornelis S, Guisez Y, Van der Heyden J, Tavernier
J 1995. Interleukin-5 and its receptor: a drug target for eosinophilia
associated with chronic allergic disease. J Leuk Biol 57:
813-819.
Devos R, Guisez Y, Plaetinck G, Cornelis S, Tavernier J, Van der Heyden
J, Foley LH, Scheffler JE 1994. Covalent modification of the interleukin-5
receptor isothiazolones leads to inhibition of binding of interleukin-5.
Eur J Bioch 225: 635-640.
Devos R, Guisez Y, Cornelis S, Verhee A, Van der Heyden J, Manneberg M,
Lahm H-W, Fiers W, Tavernier J, Plaetnick G 1993. Recombinant soluble
human interleukin-5 (hIL-5) receptor molecules: cross-linking and
stoichiometry of binding to IL-5. J Biol Chem 268:
6581-6587.
Dubucquoi S, Janin A, Klein O, Desreumaux P, Quandalle P, Cortot A,
Capron M, Colombel JF 1995. Activated eosinophils and interleukin 5
expression in early recurrence of Crohn's disease. Gut 37:
242-246.
Dunnill MS 1960. The pathology of asthma, with special reference to
changes in the bronchial mucosa. J Clin Pathol 13: 2733.
Durham SR, Ying S, Varney VA, Jacobson MR, Sudderick RM, Mackay IS, Kay
AB, Hamid QA 1992.Cytokine messenger RNA expression for IL-3, IL-4, IL-5,
and granulocyte/macrophage-colony-stimulating factor in the nasal mucosa
after local allergen provocation: relationship to tissue eosinophilia.
J Immunol 148: 2390-2394.
Egan RW, Athwahl D, Chou CC, Emtage S, Jehn CH, Kung TT, Mauser PJ,
Murgolo N, Bodmer MW 1995. Inhibition of pulmonary eosinophilia and
hyperreactivity by antibodies to interleukin-5. Intern Arch Allergy &
Immunol 107: 321-322.
Eidelman, DH, Minshall E, Dandurand RJ, Schotman E, Song YL, Yasruel Z,
Moqbel R, Hamid Q 1996. Evidence for major basic protein immunoreactivity
and IL-5 gene activation during the late phase response in explanted
airways. Am J Respir Cell Mol Biol 15: 582-589.
Elices, MJ, Osborn L, Takadu Y, Crouse C, Luhowskyj S, Hemler ME, Lobb
RR 1990. VCAM-1 on activated endothelium interacts with the leucocyte
integrin VLA-4 at a site distinct from the VLA-4/fibronectin binding site.
Cell 60: 577-584.
Endo H, Iwamoto I, Nakajima H, Yoshida S 1993. In vitro interleukin-5
production of peripheral blood mononuclear cells is increased in patients
with asthma. Intern Arch Allergy & Immunol 101: 425-430.
Eum SY, Haile S, Lefort J, Huerre M, Vargaftig BB 1995. Eosinophil
recruitment into the respiratory epithelium following antigenic challenge
in hyper-IgE mice is accompanied by interleukin 5-dependent bronchial
hyperresponsiveness. Proc Nat Acad Sci Unit Stat Am 92:
12290-12294.
Ewart SL, Mitzner W, DiSilvestre DA, Meyers DA, Levitt RC 1996. Airway
hyperresponsiveness to acetylcholine: segregation analysis and evidence for
linkage to murine chromosome 6. Am J Resp Cell & Mol Biol 14:
487-495.
Frew AJ, Kay AB 1988. The relationship between infiltrating CD4+
lymphocytes, activated eosinophils, and the magnitude of the
allergen-induced late phase cutaneous reaction in man. J Immunol
141: 4158-4164.
Fujisawa T, Abu-Ghazaleh R, Kita H, Sanderson CJ, Gleich GJ 1990.
Regulatory effect of cytokines on eosinophil degranulation. J Immunol
144: 642-646.
Fukuda T, Nakajima H, Fukushima Y, Akutsu I, Numao T, Majima K,
Motojima S, Sato Y, Takatsu K, Makino Syeau Detection of interleukin-5
messenger RNA and interleukin-5 protein in bronchial biopsies from asthma
by nonradioactive in situ hybridization and immunohistochemistry. J
Allergy & Clin Immunol 94: 584-593.
Foster PS, Hogan SP, Ramsay AJ, Matthaei KI, Young IG 1996. Interleukin
5 deficiency abolishes eosinophilia, airways hyperreactivity, and lung
damage in a mouse asthma model. J Exp Med 183: 195-201.
Gulbenkian AR, Egan RW, Fernandez X, Jones H, Kreutner W, Kung T,
Payvandi F, Sullivan L, Zurcher JA, Watnick AS 1992. Interleukin-5
modulates eosinophil accumulation in allergic guinea pig lung. Am Rev
Resp Dis 146: 263-266.
Hagan JB, Bartemes KR, Kita H, Ottesen EA, Awadzi K, Nutman TB, Gleich
GJ 1996. Elevations in granulocyte-macrophage colony-stimulating factor
and interleukin-5 levels precede posttreatment eosinophilia in
onchocerciasis. J Infect Dis 173: 1277-1280.
Hamid Q, Boguniewicz M, Leung DY 1994. Differential in situ cytokine
gene expression in acute versus chronic atopic dermatitis. J Clin
Invest 94: 870-876.
Hamid Q, Song YL, Minshall E, Bai TR, Hegele RG, Hogg JC 1996. Small
airways inflammation in asthma. Am J Resp Crit Care Med
153.4: (A878).
Hamid Q, Azzawi M, Sun Ying, Moqbel R, Wardlaw AJ, Corrigan CJ, Bradley
B, Durham SR, Collins JV, Jeffery PK, Quint DJ, Kay AB 1991. Expression of
mRNA for interleukin5 in mucosal bronchial biopsies from asthma. J Clin
Invest 87: 15411546.
Humbert M 1996. Airways inflammation in asthma and chronic bronchitis.
Clin & Exp Allergy 26: 735-737.
Humbert M, Grant JA, Tabordabarata L, Durham SR, Pfister R, Menz G,
Barkans J, Ying S, Kay AB 1996. High-affinity Ige receptor
(Fc-epsilon-ri)-bearing cells in bronchial biopsies from atopic and
nonatopic asthma. Am J Resp & Crit Care Med 153: 1931-1937.
Humbert M. Durham SR, Ying S, Kimmitt P, Barkans J, Assoufi B, Pfister
R, Menz G, Robinson DS, Kay AB, Corrigan CJ 1996. IL-4 and IL-5 mRNA and
protein in bronchial biopsies from patients with atopic and nonatopic
asthma: Evidence against asthma being a distint immunopathological entity.
Am J Resp & Crit Care Med 154: 1497-1504.
Ihle JN 1995. Cytokine receptor signalling. Nature 377:
591-594.
Ihle JN, Witthuhn BA, Quelle FW, Yamamoto K, Silvennoinen O 1995.
Signaling through the hematopoietic cytokine receptors. Ann Rev Immunol
13: 369-398.
Iwamoto I, Tomoe S, Tomioka H, Takatsu K, Yoshida S 1992. Role of CD4+
T lymphocytes and interleukin-5 in antigen-induced eosinophil recruitment
into the site of cutaneous late-phase reaction in mice. J Leuk Biol
52: 572-578.
Iwamoto T, Takatsu K 1995. Evaluation of airway hyperreactivity in
interleukin-5 transgenic mice. Intern Arch Allergy & Immunol
108 Suppl 1: 28-30.
Jeffery PK, Wardlaw AJ, Nelson FC, Collins JV, Kay AB 1989. Bronchial
biopsies in asthma. An ultrastructural, quantitative study and correlation
with hyperreactivity. Am Rev Respir Dis 140: 17451753.
Jung T, Schauer U, Rieger C, Wagner K, Einsle K, Neumann C, Heusser C
1995. Interleukin-4 and interleukin-5 are rarely co-expressed by human T
cells. Eur J Immunol 25: 2413-2416.
Kamei T, Ozaki T, Kawaji K, Banno K, Sano T, Azuma M, Ogura T 1993.
Production of interleukin-5 and granulocyte/macrophage colony-stimulating
factor by T cells of patients with bronchial asthma in response to
Dermatophagoides farinae and its relation to eosinophil colony-stimulating
factor. Am J Resp Cell & Mol Biol 9: 378-385.
Karlen S, Dercole M, Sanderson CJ 1996. Two pathways can activate the
interleukin-5 gene and induce binding to the conserved lymphokine
element. Blood 88: 211-221.
Kay AB, Ying S, Durham SR 1995. Phenotype of cells positive for
interleukin-4 and interleukin-5 mRNA in allergic tissue reactions.
Intern Arch Allergy & Immunol 107: 208-210.
Kay AB, Ying S, Durham SR 1995. Phenotype of cells positive for
interleukin-4 and interleukin-5 mRNA in allergic tissue reactions.
Intern Arch Allergy & Immunol 107: 208-210.
Kay AB, Ying S, Varney V, Gaga M, Durham SR, Moqbel R, Wardlaw AJ,
Hamid Q 1991. Messenger RNA expression of the cytokine gene cluster,
interleukin 3 (IL-3), IL-4, IL-5, and granulocyte/macrophage
colony-stimulating factor, in allergen-induced late-phase cutaneous
reactions in atopic subjects. J Exp Med 173: 775-778.
Kelso A 1995. Th1 and Th2 subsets:paradigms lost? Immunology
today 16.8: 374-379.
Kita H, Sur S, Hunt LW, Edell ES, Weiler DA, Swanson MC, Samsel RW,
Abrams JS, Gleich GJ 1996. Cytokine production at the site of disease in
chronic eosinophilic pneumonitis. Am J Resp & Crit Care Med
153: 1437-1441.
Klink M, Cline MG, Halonen M, Burrows B 1990. Problems in defining
normal limits for serum IgE. J Allergy & Clin Immunol 85:
440-444.
Koike M, Takatsu K 1994. IL-5 and its receptor: which role do they play
in the immune response. Int Arch Allergy Immunol 104: 1-9.
Kosugi H, Nakagawa Y, Hotta T, Saito H, Miyajima A, Arai K, Yokota T
1995. Structure of the gene encoding the alpha subunit of the human
interleukin 3 receptor. Bioch & Biophys Res Comm 208:
360-367.
Krishnaswamy G, Liu MC, Su SN, Kumai M, Xiao HQ, Marsh DG, Huang SK
1993. Analysis of cytokine transcripts in the bronchoalveolar lavage cells
of patients with asthma. Am J Resp Cell & Mol Biol 9: 279-286.
Kung, TT, Stelts DM, Zurcher JA, Adams III GK, Egan RW, Kreutner W,
Watnick AS, Jones H, Chapman RW 1995. Involvement of IL-5 in a murine
model of allergic pulmonary inflammation: Prophylactic and therapeutic
effect of an anti-IL-5 antibody. Am J Resp Cell Mol Biol 13:
360-365.
Kuwano K, Bosken CH, Pare PD, Bai TR, Wiggs BR, Hogg JC 1993. Small
airways dimensions in asthma and in chronic obstructive pulmonary disease.
Am Rev Respir Dis 148: 1220-1225.
Laitinen LA, Laitinen A, Haahtela T. 1993. Airway mucosal inflammation
even in patients with newly diagnosed asthma. Am Rev Respir Dis
147: 697-704.
Laviolette M, Ferland C, Comtois JF, Champagne K, Bosse M, Boulet LP
1995. Blood eosinophil leukotriene C4 production in asthma of different
severities. Eur Resp Journal 8: 1465-1472.
Lee HJ, Matsuda I, Naito Y, Yokota T, Arai N, Arai K 1994. Signals and
nuclear factors that regulate the expression of interleukin-4 and
interleukin-5 genes in helper T cells. J Allergy & Clin Immunol
94: 594-604.
Leung DM, Martin RJ, Szefler SJ, Sher ER, Ying S, Kay AB, Hamid Q 1995.
Dysregulation of Interleukin-4, Interleukin-5, and Interferon-g gene
expression in steroid resistant asthma. J Exp Med 181: 33-40.
Limaye AP, Ottesen EA, Kumaraswami V, Abrams JS, Regunathan J,
Vijayasekaran V, Jayaraman K, Nutman TB 1993. Kinetics of serum and
cellular interleukin-5 in posttreatment eosinophilia of patients with
lymphatic filariasis. J Infect Dis 167: 1396-1400.
Lopez AF, Vadas MA, Woodcock JM, Milton SE, Lewis A, Elliott MJ, Gillis
D, Ireland R, Olwell E, Park LS 1991. Interleukin-5, interleukin-3, and
granulocyte-macrophage colony stimulating factor cross-compete for binding
to cell surface receptors on human eosinophils. J Biol Chem
266: 24741-24747.
Lopez AF, Sanderson CJ, Gamble JR, Campbell HR, Young IG, Davas MA
1988. Recombinant human interleukin-5 is a selective activator of human
eosinophil function. J Exp Med 167: 219-224.
Macklem PT, Proctor DF, Hogg JC 1970. The stability of peripheral
airways. Resp Physiol 8: 191-203.
Mahanty S, Nutman TB 1993. The biology of interleukin-5 and its
receptor. Cancer Investigation 11: 624-634.
Marini M, Avoni E, Hollemborg J, Mattoli S 1992. Cytokine mRNA profile
and cell activation in bronchoalveolar lavage fluid from nonatopic
patients with symptomatic asthma. Chest 102: 661-669.
Mauser PJ, Pitman AM, Fernandez X, Foran SK, Adams GK 3rd, Kreutner W,
Egan RW, Chapman RW 1995. Effects of an antibody to interleukin-5 in a
monkey model of asthma. Am J Resp & Crit Care Med 152: 467-472.
McFadden ER Jr, Gilbert IA 1992. Asthma. New Engl J Med 327:
19281937.
Milburn MV, Hassell AM, Lambert MH, Jordan SR, Proudfoot AE, Graber P,
Wells TN 1993. A novel dimer configuration revealed by the crystal
structure at 2.4 A resolution of human interleukin-5. Nature 363:
172-176.
Migita M, Yamaguchi N, Mita S, Higuchi S, Hitoshi Y, Yoshida Y,
Tomonaga M, Matsuda I, Tominaga A, Takatsu K 1991. Characterization of the
human IL-5 receptors on eosinophils. Cellular Immunology
133: 484-497.
Minshall E, Tsicopoulos A, Yasruel Z, Bernard A, Wallaert B, Vorng H,
Tonnel A, Hamid Q 1996. High expression of cytoline gene transcripts in
active pulmonary sarcoidosis compared to non-active pulmonary sarcodoisis
and normal controls. Am J Respir Crit Care Med 153: A868.
Miyajima A, Kitamura T, Harada N, Yokota T, Arai K 1992. Cytokine
receptors and signal transduction. Ann Rev Immunol 10:
295-331.
Matsumoto K, Schleimer RP, Saito H, Iikura Y, Bochner BS 1995.
Induction of apoptosis in human eosinophils by anti-Fas antibody treatment
in vitro. Blood 86: 1437-1443.
Miyajima A, Mui AL, Ogorochi T, Sakamaki K 1993. Receptors for
granulocyte-macrophage colony-stimulating factor, interleukin-3, and
interleukin-5. Blood 82: 1960-1974.
Miyajima A, Mui AL, Ogorochi T, Sakamaki K 1993. Receptors for
granulocyte-macrophage colony-stimulating factor, interleukin-3, and
interleukin-5. Blood 82: 1960-1974.
Modlin RL, Nutman TB 1993. Type 2 cytokines and negative immune
regulation in human infections. Curr Opin Immunol 5: 511-517.
Moqbel R, Barkans J, Bradley BL, Durham SR, Kay AB 1992. Application of
monoclonal antibodies against major basic protein (BMK13) and eosinophil
cationic protein (EG1 and EG2) for quantifying eosinophils in brochial
biopsies from atopic asthma. Clin Exp Allergy 22: 265273.
Moser R, Fehr J, Bruijnzeel PLB 1992. IL-4 controls the selective
endothelium driven transmigration of eosinophils from allergic
individuals. J Immunol 149: 1432-1438.
Motojima S, Akutsu I, Fukuda T, Makino S, Takatsu K 1993. Clinical
significance of measuring levels of sputum and serum ECP and serum IL-5 in
bronchial asthma. Allergy 48 (17 Suppl): 98-106.
Mui AL, Wakao H, Harada N, O'Farrell AM, Miyajima A 1995.
Interleukin-3, granulocyte-macrophage colony-stimulating factor, and
interleukin-5 transduce signals through two forms of STAT5. J Leuk Biol
57: 799-803.
Murata, Y, Takaki S, Migita S, Kikuchi Y, Tominaga A, Takatsu K 1992.
Molecular cloning and expression of the human interleukin-5 receptor. J
Exp Med 175: 341-351.
Nagai H, Yamaguchi S, Inagaki N, Tsuruoka N, Hitoshi Y, Takatsu K 1993.
Effect of anti-IL-5 monoclonal antibody on allergic bronchial eosinophilia
and airway hyperresponsiveness in mice. Life Sciences. 53:
Pl 243-247.
Nakajima H, Iwamoto I, Tomoe S, Matsumura R, Tomioka H, Takatsu K,
Yoshida S 1992. CD4+ T-lymphocytes and interleukin-5 mediate
antigen-induced eosinophil infiltration into the mouse trachea. Am Rev
Respir Dis 146: 374-377.
Naora H, Young IG 1995. Comparison of the mechanisms regulating IL-5,
IL-4, and three other lymphokine genes in the Th2 clone D10.G4.1. Exp
Hematol 23: 597-602.
Nicola NA 1989. Hemopoietic cell growth factors and their receptors.
Ann Rev Biochem 58: 45-77.
Noelle R, Snow EC 1992. T helper cells. Curr Opin Immunol 4:
333-337.
Ogawa M 1994. Hematopoiesis. J Allergy & Clin Immunol 94:
645-650.
Ohnishi T, Sur S, Collins DS, Fish JE, Gleich GJ, Peters SP 1993.
Eosinophil survival activity identified as interleukin-5 is associated
with eosinophil recruitment and degranulation and lung injury twenty-four
hours after segmental antigen lung challenge. J Allergy & Clin
Immunol 92: 607-615.
Okudaira H, Mori A, Suko M, Etoh T, Nakagawa H, Ito K 1995. Enhanced
production and gene expression of interleukin-5 in patients with bronchial
asthma: possible management of atopic diseases by down-regulation of
interleukin-5 gene transcription. Internat Arch Allergy & Immunol
107: 255-258.
Olivenstein R, Renzi PM, Yang JP, Rossi P, Laberge S, Waserman S,
Martin JG 1993. Depletion of OX-8 lymphocytes from the blood and airways
using monoclonal antibodies enhances the late airway response in rats.
J Clin Invest 92: 1477-1482.
Pazdrak K, Schreiber D, Forsythe P, Justement L, Alam R 1995. The
intracellular signal transduction mechanism of interleukin 5 in
eosinophils: the involvement of lyn tyrosine kinase and the
Ras-Raf-1-MEK-microtubule-associated protein kinase pathway. J Exp Med
181: 1827-1834.
Pene J, Rousset F, Briere F, Chretien I, Wideman J, Bonnefoy JY, De
Vries JE 1988. Interleukin 5 enhances interleukin 4-induced IgE production
by normal human B cells. The role of soluble CD23 antigen. Eur J
Immunol 18: 929-935.
Plaut M, Pierce JH, Watson CJ, Hanley-Hyde J, Nordan RP, Paul WE 1989.
Mast cell lines produce lymphokines in response to cross-linkage of Fc
epsilon RI or to calcium ionophores. Nature 339: 64-67.
Pradalier A 1993. Late-phase reaction in asthma: basic mechanisms.
Internat Arch Allergy & Immunol 101: 322-325.
Purkerson JM, Isakson PC 1992. Interleukin 5 (IL-5) provides a signal
that is required in addition to IL-4 for isotype switching to
immunoglobulin (Ig) G1 and IgE. J Exp Med 175: 973-982.
Quan SF, Sedgwick JB, Nelson MV, Busse WW 1993. Corticosteroid
resistance in eosinophilic gastritis-relation to in vitro
eosinophil survival and interleukin 5. Annals of Allergy 70:
256-260.
Renzi PM, Du T, Sapienza S, Wang NS, Martin JG 1991. Acute effects of
interleukin-2 on lung mechanics and airway responsiveness in rats. Am
Rev Respir Dis 143: 380-385.
Renzi PM, Sapienza S, Du T, Wang NS, Martin JG 1991. Lymphokine-induced
airway hyperresponsiveness in the rat. Am Rev Respir Dis
143: 375-379.
Renzi PM, Olivenstein R, Martin JG 1993. Inflammatory cell populations
in the airways and parenchyma after antigen challenge in the rat. Am
Rev Respir Dis 147: 967-974.
Renzi PM, Yang JP, Yasruel Z, Al Assaad S, Hamid Q 1996. Cytokine
expression in the presence or absence of late airway responses after
antigen challenge of sensitized rats. Am J Respir Cell Mol Biol
15: 367-373.
Robinson DS, Hamid Q, Bentley A, Ying S, Kay AB, Durham SR 1993.
Activation of CD4+ T cells increased Th2-type cytokine mRNA expression and
eosinophil recruitment in bronchoalveolar lavage after allergen inhalation
challenge in patients with atopic asthma. J Allergy Clin Immunol
92: 313-324.
Robinson D, Hamid Q, Ying S, Bentley A, Assoufi B, Durham S, Kay AB
1993. Prednisone treatment in asthma is associated with modulation of
bronchoalveolar lavage cell Interleukin-4, Interleukin-5, and Interferon-g
cytokine gene expression. Am Rev Respir Dis 148: 401-406.
Robinson DS, Ying S., Bentley AM, Meng Q, North J, Durham SR, Kay AB,
Hamid Q 1993. Relationships among numbers of bronchoalveolar lavage cells
expressing messenger ribonucleic acid for cytokines, asthma symptoms, and
airway methacholine responsiveness in atopic asthma. J Allergy Clin
Immunol 92: 397403.
Robinson DS, Hamid Q, Ying S, Tsicopoulos A, Barkans J, Bentley AM,
Corrigan C, Durham SR, Kay AB 1992.Predominant Th2-like bronchoalveolar
T-lymphocyte population in atopic asthma. N Engl J Med 326:
298-304.
Rothenberg ME, Ownbey R, Mehlhop PD, Loiselle PM, Vanderijn M,
Bonventre JV, Oettgen HC, Leder P, Luster AD 1996. Eotaxin triggers
eosinophil-selective chemotaxis and calcium flux via a distinct receptor
and induces pulmonary eosinophilia in the presence of interleukin 5 in
mice. Molecular Medicine 2: 334-348.
Sanderson CJ 1992. Pharmacological implications of interleukin-5 in the
control of eosinophilia. Advances in Pharmacol 23: 163-177.
Sanderson CJ 1992. Interleukin-5, eosinophils, and disease. Blood
79: 3101-3109.
Sakamaki K, Miyajima I, Kitamura T, Miyajima A 1992. Critical
cytoplasmic domains of the common beta subunit of the human GM-CSF, IL-3
and IL-5 receptors for growth signal transduction and tyrosine
phosphorylation. EMBO Journal 11: 3541-3549.
Satoh T, Sun L, Li MS, Spry CJ 1994. Interleukin-5 mRNA levels in blood
and bone marrow mononuclear cells from patients with the idiopathic
hypereosinophilic syndrome. Immunol 83: 308-812.
Schrezenmeier H, Thome SD, Tewald F, Fleischer B, Raghavachar A 1993.
Interleukin-5 is the predominant eosinophilopoietin produced by cloned T
lymphocytes in hypereosinophilic syndrome. Experimental Hematol
21: 358-365.
Sedgwick JB, Quan SF, Calhoun WJ, Busse WW 1995. Effect of
interleukin-5 and granulocyte-macrophage colony stimulating factor on
in vitro eosinophil function: comparison with airway eosinophils.
J Allergy & Clin Immunol 96: 375-385.
Sewell WA, Mu HH1996. Dissociation of production of interleukin-4 and
interleukin-5. Immunol & Cell Biol 74: 274-277.
Sur S, Gleich GJ, Swanson MC, Bartemes KR, Broide DH 1995. Eosinophilic
inflammation is associated with elevation of interleukin-5 in the airways
of patients with spontaneous symptomatic asthma. J Allergy & Clin
Immunol 96: 661-668.
Taha R, Kotsimbos T, Song YL, Schotman E, Menzies R, Hamid Q 1997.
IFN-g and IL-12 are increased in active vs. non-active pulmonary
tuberculosis. Am J. Resp& Crit Care Med 155: 1135-1139.
Takagi M, Hara T, Ichihara M, Takatsu K, Miyajima A 1995. Multi-colony
stimulating activity of interleukin 5 (IL-5) on hematopoietic progenitors
from transgenic mice that express IL-5 receptor alpha subunit
constitutively. J Exp Med 181: 889-899.
Takai S, Yamada K, Hirayama N, Miyajima A, Taniyama T 1994. Mapping of
the human gene encoding the mutual signal-transducing subunit (beta-chain)
of granulocyte-macrophage colony-stimulating factor (GM-CSF),
interleukin-3 (IL-3), and interleukin-5 (IL-5) receptor complexes to
chromosome 22q13.1. Human Genetics 93: 198-200.
Takaki S, Murata Y, Kitamura T, Miyajima A, Tominaga A, Takatsu K 1993.
Reconstitution of the functional receptors for murine and human
interleukin 5. J Exp Med 177: 1523-1529.
Tavernier J, Tuypens T, Plaetinck G, Verhee A, Fiers W, Devos R 1992.
Molecular basis of the membrane-anchored and two soluble isoforms of the
human interleukin 5 receptor subunit. Proc Natl Acad Sci 89:
7041-7045 .
Tavernier J, Devos R, Cornelis S, Tuypens T, Van der Heyden J, Fiers W,
Plaetinck G 1991. A human high affinity interleukin-5 receptor (IL-5) is
composed of an IL-5-specific a chain and a b chain shared with the
receptor for GM-CSF. Cell 66: 1175-1184.
Till S, Li B, Durham S, Humbert M, Assoufi B, Huston D, Dickason R,
Jeannin P, Kay AB, Corrigan C 1995. Secretion of the eosinophil-active
cytokines interleukin-5, granulocyte/macrophage colony-stimulating factor
and interleukin-3 by bronchoalveolar lavage CD4+ and CD8+ T cell lines in
atopic asthmatics, and atopic and non-atopic controls. Eur J Immunol
25: 2727-2731.
Tuypens T, Plaetinck G, Baker E, Sutherland G, Brusselle G, Fiers W,
Devos R, Tavernier J 1992. Organization and chromosomal localization of
the human interleukin 5 receptor alpha-chain gene. Euro Cytokine
Network 3: 451-459.
Van der Bruggen T, Caldenhoven E, Kanters D, Coffer P, Raaijmakers JA,
Lammers JW, Koenderman L 1995. Interleukin-5 signaling in human
eosinophils involves JAK2 tyrosine kinase and Stat1 alpha. Blood
85: 1442-1448.
Van Oosterhout AJ, Ladenius AR, Savelkoul HF, Van Ark I, Delsman KC,
Nijkamp FP 1993. Effect of anti-IL-5 and IL-5 on airway hyperreactivity
and eosinophils in guinea pigs. Am Rev Respir Dis 147: 548-552.
Van Oosterhout AJ, Fattah D, Van Ark I, Hofman G, Buckley TL, Nijkamp
FP 1995. Eosinophil infiltration precedes development of airway
hyperreactivity and mucosal exudation after intranasal administration of
interleukin-5 to mice. J Allergy & Clinical Immunol 96:
104-112.
Van Straaten JF, Dokter WH, Stulp BK, Vellenga E 1994. The regulation
of interleukin 5 and interleukin 3 gene expression in human T cells.
Cytokine 6: 229-234.
Walker C, Bauer W, Braun RK, Menz G, Braun P, Schwarz F, Hansel TT,
Villiger B 1994. Activated T cells and cytokines in bronchoalveolar
lavages from patients with various lung diseases associated with
eosinophilia. Am J Respir Critical Care Med 150: 1038-1048.
Walker C, Bode E, Boer L, Hansel TT, Blaser K, Virchow JC Jr 1992.
Allergic and nonallergic asthmatics have distinct patterns of T-cell
activation and cytokine production in peripheral blood and bronchoalveolar
lavage. Am Rev Respir Dis 146: 109-115.
Walker C, Kaegi MK, Braun P, Blaser K 1991. Activated T cells and
eosinophilia in bronchoalveolar lavages from subjects with asthma
correlated with disease severity. J Allergy Clin Immunol 88:
935942.
Watanabe A, Rossi P, Renzi PM, Xu LJ, Guttmann RD, Martin JG 1995.
Adoptive transfer of allergic airway responses with sensitized lymphocytes
in BN rats. Am J Respir & Critical Care Med 152: 64-70.
Watanabe A, Mishima H, Renzi PM, Xu LJ, Hamid Q, Martin JG 1995.
Transfer of allergic airway responses with antigen-primed CD4+ but not
CD8+ T cells in brown Norway rats. J Clin Invest 96: 1303-1310.
Watanabe A, Mishima H, Kotsimbos ATC, Renzi PM, Martin JG, Hamid Q
1997. Adoptively transferred late allergic airway responses are associated
with Th2 type cytokines in the rat. Am J Respir Crit Care Med
16: 69-74.
Weller PF 1992. Cytokine regulation of eosinophil function. Clinical
Immunol & Immunopathol 62: S55-59.
Wiggs BR, Bosken C, Pare PD, Hogg JC 1992. A model of airway narrowing
in asthma and in chronic obstructive pulmonary disease. Am Rev Respir
Dis145: 1251-1258.
Yamagata T, Nishida J, Sakai R, Tanaka T, Honda H, Hirano N, Mano H,
Yazaki Y, Hirai H 1995. Of the GATA-binding proteins, only GATA-4
selectively regulates the human interleukin-5 gene promoter in
interleukin-5-producing cells which express multiple GATA-binding
proteins. Molecular & Cellular Biology 15: 3830-3839.
Yamaguchi S, Nagai H, Tanaka H, Tsujimoto M, Tsuruoka N. 1994. Time
course study for antigen-induced airway hyperreactivity and the effect of
soluble IL-5 receptor. Life Sciences 54: 471-475.
Yamaguchi, Y, Hayashi Y, Sugama Y, Miura Y, Kasahara T, Kitamura S,
Torisu M, Mita S, Tominaga A, Takatsu K, Suda T 1988. Highly purified
murine interleukin-5 (IL-5) stimulates eosinophil function and prolongs
in vitro survival. J Exp Med 167:1737-1742.
Yamaguchi N, Hitoshi Y, Takaki S, Murata Y, Migita M, Kamiya T,
Minowada J, Tominaga A, Takatsu K 1991. Murine interleukin 5 receptor
isolated by immunoaffinity chromatography: comparison of determined
N-terminal sequence and deduced primary sequence from cDNA and implication
of a role of the intracytoplasmic domain. Internat Immunol 3:
889-98.
Yang LP, Byun DG, Demeure CE, Vezzio N, Delespesse G 1995. Default
development of cloned human naive CD4 T cells into interleukin-4- and
interleukin-5- producing effector cells. Eur J Immunol 25:
3517-3520.
Yasruel Z, Humbert M, Kotsimbos ATC, Ploysongsang Y, Minshall E, Durham
S, Pfister R, Menz G, Tavernier J, Kay AB, Hamid Q 1997. Expression of
membrane-bound and soluble interleukin-5 alpha receptor mRNA in the
bronchial mucosa of atopic & non-atopic asthmatics. Am J Resp Crit Care
Med 155: 1413-1418.
Ying S, Durham SR, Corrigan CJ, Hamid Q, Kay AB 1995. Phenotype of
cells expressing mRNA for TH2-type (interleukin 4 and interleukin 5) and
TH1-type (interleukin 2 and interferon gamma) cytokines in bronchoalveolar
lavage and bronchial biopsies from atopic asthmatic and normal control
subjects. Am J Respir Cell & Mol Biol 12: 477-487.
Ying S, Durham SR, Barkans J, Masuyama K, Jacobson M, Rak S, Lowhagen
O, Moqbel R, Kay AB, Hamid Q 1993. T cells are the principal source of
interleukin5 mRNA in allergeninduced rhinitis. Am J Respir Cell Mol
Biol 9: 356360.
Ying S, Durham SR, Barkans J, Masuyama K, Jacobson M, Rak S, Lowhagen
O, Moqbel R, Kay AB, Hamid QA 1993. T cells are the principal source of
interleukin-5 mRNA in allergen-induced rhinitis. Am J Respir Cell Mol
Biol 9: 356-360.
Zanders ED 1994. Interleukin-5 receptor alpha chain mRNA is
downregulated by transforming growth factor-b1. Soluble or
membrane-anchored? Eur Cytokine Network 5: 419-422.
Zon LI, Yamaguchi Y, Yee K, Albee EA, Kimura A, Bennett JC, Orkin SH,
Ackerman SJ 1993. Expression of mRNA for the GATA-binding proteins in
human eosinophils and basophils: potential role in gene transcription.
Blood 81: 3234-3241.
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc97172b.jpg]
[oc97172g.jpg]
[oc97172d.jpg]
[oc97172f.jpg]
[oc97172a.jpg]
[oc97172e.jpg]
[oc97172c.jpg]
|

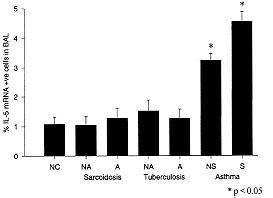{kind=link}
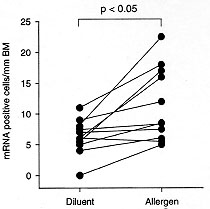{kind=link}
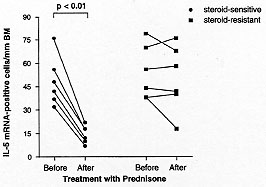{kind=link}
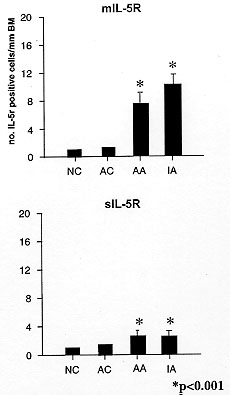{kind=link}
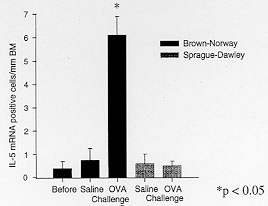{kind=link}
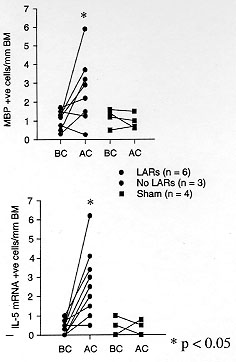{kind=link}
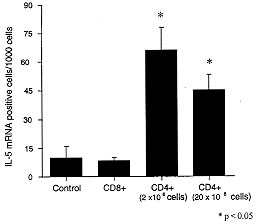{kind=link}