
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 92, Num. s2, 1997, pp. 109-114
|
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92 (Suppl.II), 1997, pp.109-114
Cytokines in the Modulation of Eosinophilia
Lucia H Faccioli/^+, BB Vargaftig*, Alexandra I Medeiros, Adriana
Malheiros
Departamento de Parasitologia, Microbiologia e Imunologia, Faculdade de
Medicina de Ribeirao Preto, Universidade de Sao Paulo, Av. Bandeirantes
3900, 14049-900 Ribeirao Preto, SP, Brasil
* Unite de Pharmacologie Cellulaire, Unite Associee Institut
Pasteur/INSERM, Paris, France
^+Corresponding author. Fax: + 55-16-635.6631. E-mail: faccioli@fmrp.usp.br
Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97175
Sizes of Files:
Text: 21.7K
Graphics: Line drawings and photographs (jpg) - 87.5K
Tables (jpg) - 60.4K
In this review we discuss our recently results showing interleukin 5
(IL-5) involvement in eosinophil migration and in the maintenance of
eosinophilia in blood, bone marrow, lung and peritoneal cavity, in a
visceral larva migrans syndrome model using guinea-pigs infected with
Toxocara canis. We also describe the sequential release of TNF-alpha
and IL-8 during the course of infection, and the interaction between these
cytokines and IL-5 during infection. Finally we propose a new biological
role for IL-5, at least in our model, as a modulator of IL-8 release and
secretion.
Key words: eosinophils - helminth Parasites - inflammation -
cytokines
Toxocara canis is an intestinal parasite of dogs, and is the most
common aetiologic agent of visceral larva migrans syndrome (VLMS) (Beaver
et al. 1952). Tissue-migrating larvae of this parasite induce intense
eosinophilia which reaches more than 90% of total leukocyte counts (Beaver
et al. 1952). Although several investigators have suggested a direct
correlation between eosinophilia and interleukin-5 (IL-5) in human
helminthic infection (Limaye et al. 1990, Steel & Nutman 1993) and in
experimental animal models (Yamaguchi et al. 1990, Parson et al. 1993), the
mechanisms involved in blood and tissue eosinophilia in this model of
helminthic infection remain unclear. The release of other
eosinophil-related cytokines such as IL-8 and TNF-alpha and their interaction
with IL-5 are currently being studied.
Widespread eosinophilia and eosinophil migration in VLMS is IL-5
dependent
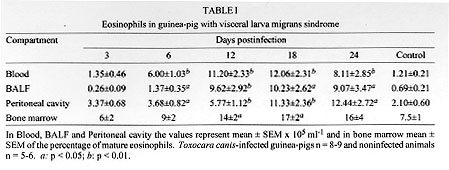
Guinea-pigs infected orally with T. canis eggs showed widespread
eosinophilia with a time-dependent increase of eosinophils in all
compartments studied (Table
I). In blood and bronchoalveolar
lavage fluid (BALF) the number of eosinophils was significantly increased
at 6 days post-infection reaching more than 90% of the total cell counts in
BALF but decreased by day 24. In contrast to blood and BALF, the number of
eosinophils in the peritoneal cavity increased significantly only at day 12
post-infection and increased progressively until day 24. The percentage of
eosinophils in some animals reached 55% at the peak of infection (Faccioli
et al. 1996).
The development of eosinophilia in guinea-pigs infected with T.
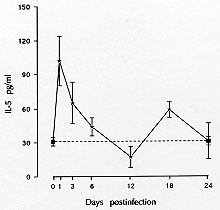
canis is accompanied by the release of two peaks of serum IL-5. The
highest occurs soon after the stimulus, i.e., 1 day after infection, and
the second occurs 18 days later (Fig. 1). Since IL-5 release
correlated with the percentage of larvae recovered from the liver of
guinea-pigs (Faccioli et al. 1996), led us to suggest that the eosinophilia
against helminth larvae is initiated by the release of IL-5 when the
parasites migrate from the intestine to the liver by stimulation of a
specific cell population. The cytokine pattern that develops at this early
stage probably is a T-cell independent pathway which may also influence the
subsequent T-cell differentiation into Th2 type, which may be responsible
for the second peak.
Fig. 1: IL-5
concentration in serum of Toxocara
canis-infected guinea-pigs (n = 3-5). Basal IL-5
concentrations in noninfected animals were 31 +/- 4 pg/ml (doted line).
IL-5 levels in the serum of guinea-pigs were measured by an enzyme-liked
immunosorbent assay (Faccioli et al. 1996).
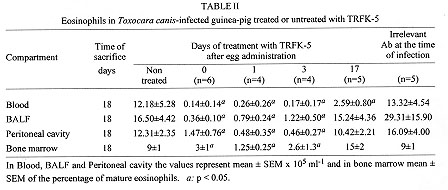
In our study, i.p. administration of TRFK-5, a monoclonal antibody (mAb)
against IL-5, at the time of egg administration or one or three days later,
drastically reduced the number of eosinophils in blood, BALF, peritoneal
cavity and bone marrow by 18 days after infection (Table II).
TRFK-5 administered 17 days after infection, and the animals killed 24 hr
later, significantly inhibited the number of circulating eosinophils but
was accompanied by an increase of mature eosinophils in bone marrow. The
inhibition of circulating eosinophil numbers by different treatments with
mAb, even when the antibody was given at the peak of blood eosinophilia,
suggests that IL-5, apart from being required for terminal differentiation
of eosinophils in bone marrow (Rennick et al. 1990), may also drive
eosinophils from the bone marrow to blood and then to tissues (probably by
up-regulating VLA-4 expression in eosinophils). Thus, even though the
increase of serum IL-5 level shows only two peaks followed by levels close
to the control during the rest of infection, the maintenance of the basal
level of serum IL-5 appears to be essential for eosinophilia and to drive
eosinophils during the infection.
Serum TNFa and IL-8 levels in VLMS
Since little is known about the presence of other cytokines related to
eosinophil recruitment in the model of T. canis infection, we
determined the release of two relevant cytokines related to eosinophils
migration, TNF-alpha (Weg et al. 1995, Lukacs et al. 1995a) and IL-8 (Collins
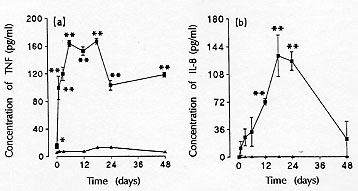
et al. 1993, Sehmi et al. 1993). TNF-alpha levels increased very early in the
serum of infected guinea-pigs, reaching levels 87% above those of
noninfected animals 4 hr after egg inoculation and remaining above control
levels up to 48 days post-infection (Fig. 2a). T. canis
larvae persist in different organs for long periods of time in infected
animals (Kayes & Oaks 1976) and release excretory-secretory antigens during
migration (Parsons et al. 1986). The prolonged and sustained TNF-u activity
in blood may be explained either by the persistent presence of larvae in
the tissues and/or by the release of those excretory-secretory antigens.
TNF-alpha is involved in eosinophil recruitment (Weg et al. 1995, Lukacs et al.
1995a) and has also been reported to occur in diseases presenting elevated
number of eosinophils, such as late (Gosset et al. 1991) and early (1 to 8
hr) asthmatic reactions during airway inflammation (Lukacs et al. 1995a).
Finally, bronchoalveolar leukocytes from patients with bronchial asthma
secrete high levels of TNF (Cembrzyanska-Novak et al. 1993).
Figure 2 : serum
TNF-u (a), and IL-8 (b) levels in
Toxocara canis-infected (n) guinea-pigs sacrificed at different
times post-infection (n = 4-5 per day). Asterisks indicate a
significant difference between infected and non-infected animals (s). * p<
0.05 and ** p< 0.01. Serum cytokines were measured using human ELISA kits.
The IL-8 serum levels showed a similar profile to that of blood
eosinophilia, increasing significantly only between 6 and 12 days
post-infection and peaking between days 18 and 24 (Fig. 2b).
These increases occurred at the same time as the increase in circulating
eosinophil numbers, and the decrease of IL-8 in serum was followed by a
reduction in blood eosinophil counts. This suggests that IL-8 contributed
to eosinophil recruitment. IL-8 has been described as an eosinophil
chemoat-tractant in vivo and in vitro (Collins et al. 1993,
Sehmi et al. 1993, Erger & Casale 1995). However, it appears that
IL-8 is only able to induce eosinophil migration after being primed by
IL-5 (Moser et al. 1992, Warringa et al. 1992, Sehmi et al. 1993). Based on
these data and on the data presented here, we suggest that IL-8 requires
pre-priming with IL-5 to induce eosinophil recruitment, in vivo.
Thus, despite the release of high amounts of IL-8 in this model, the
presence of IL-5 was essential for eosinophil migration, as demonstrated in
antigen-challenged guinea-pigs (Coeffier et al. 1994) and in vitro
(Moser et al. 1992, Warringa et al. 1992, Sehmi et al. 1993). However, the
exact mechanisms involved in this process are not yet completely
understood, and further studies using anti-IL-8 Ab will be essential to
determine the contribution of IL-8 to the eosinophil recruitment and lung
inflammation (Faccioli et al. 1996) occurring in this model. The
observation that serum IL-8 level started to increase only after serum TNF
release suggests that TNF may be involved in the induction of IL-8 release
in this model. Indeed, several reports have shown that TNF is a potent
inductor of IL-8 (Kunkel et al. 1990, Kwon et al. 1994). IL-8 is present
after TNF therapy in patients with chronic hepatitis (Sheron & William
1992) and is released in vitro from pulmonary smooth muscle and
endothelial cells stimulated with TNF (Lukacs et al. 1995b).
IL-5 modulates IL-8 synthesis and release during helminthic infection
and in supernatants of LPS-stimulated guinea-pig adherent peritoneal cells
During the course of infection we observed a sequential release of TNF-alpha
and IL-8 in connection with IL-5 release (Faccioli et al., 1996). We also
examined the relationship between IL-5 levels and IL-8 and TNF-alpha release
in vivo by inducing IL-5 depletion with anti-IL-5 mAb.
Intraperitoneal injection of anti-IL-5 mAb (2 mg/animals) into T.
canis-infected guinea-pigs only at the time of egg administration of
several doses (0.3 mg/animal on days 0, 1, 3 and 17 post-infection)
inhibited blood eosinophil counts by 95% to 100 %, as shown in Fig. 3a. When the
animals were treated with a single
dose of anti-IL-5 mAb, a 159% increase in IL-8 was observed in serum (Fig. 3c). Moreover, when
infected animals were treated with
several doses, eosinophilia was also suppressed and serum IL-8 levels were
increased by 216% (Fig. 3c). By contrast, no alteration in serum TNF
concentration was observed regardless of the treatment (Fig.
3b). Infected animals treated with irrelevant mAb showed no
significant differences in serum IL-8 and TNF levels (Faccioli et al.
unpublished data).
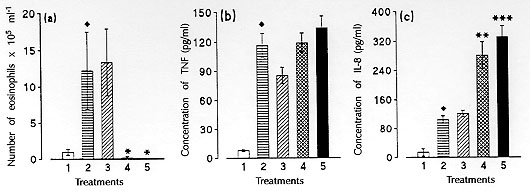
Figure 3: blood
eosinophil counts (a), serum TNF (b),
and IL-8 (c) levels in untreated Toxocara canis-infected guinea-pigs
(column 2), and in infected animals treated with irrelevant Ab (column 3),
or treated with anti-IL-5 Ab once at the time of infection (column 4) or
with several doses (0.3 mg/animal on days 0, 1, 3 and 17 post-infection)
(column 5). Animals were sacrificed 18 days postinfection. Diamonds
indicate a significant difference between untreated infected animals (n
= 5-6) and non-infected controls (column 1, n = 6). u p< 0.05.
Asterisks indicate a significant difference between untreated infected
animals (n = 5-6) and infected animals treated with anti-IL-5 Ab
(n =7). ** p< 0.01 and *** p< 0.001 (Faccioli et al. unpublished
data).
To corroborate the results obtained in vivo, we carried out
experiments in vitro to examine the effect of recombinant IL-5
(rIL-5) on IL-8 release in supernatants of LPS-stimulated adherent
peritoneal cells and on IL-8 and IL-5 mRNA expression. IL-8 was determined
in supernatants of LPS-stimulated guinea-pig adherent peritoneal cells
pre-incubated or not with rIL-5 (Fig. 4). LPS-induced IL-8
release was inhibited (64% to 66%), by rIL-5 addition. Moreover, when rIL-5
was added to the cells before LPS, a marked inhibition of IL-8 release and
IL-8 mRNA expression occurred, which was prevented by anti-IL-5 mAb
treatment, indicating specificity (Faccioli et al. unpublished data)
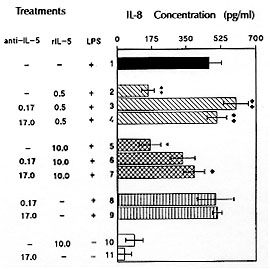
Fig. 4: inhibition
of IL-8 generation into supernatants
of LPS-stimulated (500 ng/ml) guinea-pig adherent peritoneal cells
pre-treated with 0.5 ng/ml or 10 ng/ml of rIL-5. Data are presented as
means +/-SEM (n = 4). Asterisks indicate differences between the
amount of IL-8 released by LPS-stimulated adherent peritoneal cells
pre-incubated without or with rIL-5. * p< 0.05 and ** p< 0.01. Diamonds
indicate differences between the amount of IL-8 released by LPS-stimulated
adherent peritoneal cells pre-treated with rIL-5 in the absence or presence
of anti-IL-5 Ab (0.17 ng/ml or 17 ng/ml). u p< 0.05 and uu p< 0.01
(Faccioli et al. unpublished data).
These data suggest a regulatory role of IL-5 acting on IL-8 synthesis and
perhaps on secretion, in vivo. Thus, the inhibitory effects of IL-5
on IL-8 expression and synthesis may represent an endogenous
down-regulating mechanism for eosinophil inflammation. It is possible that
IL-5 is required to prevent IL-8 over production and release. Moreover, as
discussed above, we suggest that serum IL-8, in the presence of IL-5, acts
as an eosinophil chemoattractant. However, since IL-5 is decreased during
the course of infection an increase in serum IL-8 occurs which, above a
certain level, may act as a modulator of eosinophil release from bone
marrow and tissue eosinophil migration. Thus, it is possible that the
increase in serum IL-8 levels in T. canis-infected guinea-pigs after
anti-IL-5 mAb treatment may contribute to the inhibition of eosinophilia.
IL-8, which is a potent chemotactic factor for neutrophils has been shown
to inhibit migration to the skin when administered at high concentrations
by the i.v. route (Hechtman et al. 1991).
The present results describe a new function of IL-5 as a modulator of IL-8
synthesis and secretion and may help in understanding and ultimate control
of diseases associated with eosinophilia and high IgE levels (Nutman et al.
1989). IL-8 has been shown to selectively inhibit IgE production (Kimata et
al. 1992, 1995) and thus may be involved in the control of serum IgE levels
in vivo. Production of high amounts of IL-5 in such diseases may
down regulate IL-8 levels and consequently contribute to the maintenance of
high IgE levels during infection.
In summary, our results demonstrate that in T. canis-induced
eosinophilia there is a sequential release of TNF-alpha and IL-8 which occurr
in parallel to or after serum IL-5 release. Thus, in this model, IL-5
appears to be the main factor involved in both induction of eosinophilia
and eosinophil migration. Also, IL-5 may modulate eosinophilic inflammation
by down-regulating IL-8 synthesis and secretion.
REFERENCES
Beaver P, Snyder H, Carrera G, Dent J, Lafferty J 1952. Chronic
eosinophilia due to visceral larva migrans. Pediatrics 9: 7-19.
Cöeffier E, Joseph D, Vargaftig BB 1994. Role of interleukin-5 in
enhanced migration of eosinophils from airways of immunized guinea-pigs.
Br J Pharmacol 113: 749-756.
Cembrzyanska-Novak M, Szklarz E, Inglot AD, Teodorczyk-Injeyan JA 1993.
Elevated release of tumor necorsis factor-alpha and interferon-gamma by
bronchoalveolar leukocytes from patients with bronchial asthma. Am Rev
Respir Dis 147: 291-295.
Collins P, Weg VB, Faccioli LH, Watson ML, Moqbel R, Williams TJ 1993.
Eosinophil accumulation induced by human interleukin-8 in the guinea-pig
in vivo. Immunology 79: 312-318.
Erger RA, Casale TB 1995. Interleukin-8 is a potent mediator of
eosinophil chemotaxis through endothelium and epithelium. Am J Physiol
268: L117-L122.
Faccioli LH, Mokwa VF, Silva CL, Rocha GM, Araujo JI, Nahori MA,
Vargaftig BB 1996. IL-5 drives eosinophils from bone marrow to blood and
tissues in a guinea-pig model of visceral larva migrans syndrome.
Mediators of Inflammation 5: 24-31.
Gosset P, Tsicopoulos A, Wallaert B, Vannimerus C, Joseph M, Tonnel
A-B, Capron A 1991. Increased secretion of tumor necrosis factor a and
interleukin 6 by alveolar macrophage consecutive to the development of the
late asthmatic reaction. J Allergy Clin Immunol 88: 561-571.
Hechtman DH, Cybulsky MI, Fuchs HJ, Baker JB, Gimbrone MA 1991.
Intravascular IL-8. Inhibitor of polymorphonuclear leukocyte accumulation
at sites of acute inflammation. J Immunol 147: 883-892.
Kayes SG, Oaks JA 1976. Effect of inoculum size and length of infection
on the distribution of Toxocara canis larvae in the mouse. Am J
Trop Med Hyg 25: 573-580.
Kimata H, Yoshida A, Ishioka C, Lindley I, Mikawa H 1992. Interlukin 8
(IL-8) selectively inhibits immunoglobulin E production induced by IL-4 in
human B cells. J Exp Med 176: 1227-1231.
Kimata H, Lindley I, Furusho K 1995. Selective inhibition of
spontaneous IgE and IgG4 production by interleukin-8 in atopic patients.
Blood 85: 3191-3198.
Kunkel SL, Strieter RM, Chensue SW, Basha M, Standiford T, Ham J,
Remick DJ 1990. Tumor necrosis factor-alpha, interleukin-8 and chemotactic
cytokines. Prog Clin Biol Res 349: 433-444.
Kwon OJ, Au BT, Collins PD, Adcock IM, Mak JC, Robbins RR, Fan Chung K,
Barnes PJ 1994. Tumor necrosis factor-induced interleukin-8 expression in
cultured human airway epithelial cell. Am J Physiol 267: L398-L405.
Limaye AP, Abrams JS, Silver JE, Ottesen EA, Nutman TB 1990. Regulation
of parasite-induced eosinophilia: Selectively increased interleukin 5
production in helminthic-infected patients. J Exp Med 172: 399-402.
Lukacs NW, Strieter RM, Chensue SW, Windmer M, Kunkel SL 1995a. TNFa
mediates recruitment of neutrophils and eosinophils during airway
inflammation. J Immunol 154: 5411-5417.
Lukacs NW, Kunkel SL, Allen R, Evanoff HL, Shaklee CL, Sherman JS,
Burdick MD, Strieter RM 1995b. Stimulus and cell-specific expression of
C-X-C and C-C chemokines by pulmonary stromal cell populations. Lungs
Cells Mol Physiol 12: L856-L861.
Moser R, Fehr J, Olgiati L, Bruijnzeel PL 1992. Migration of primed
human eosinophils across cytokine-activated endothelial cell monolayers.
Blood 79: 2937-2945.
Nutman TB, Ottesen EA, Cohen SG 1989. The eosinophil, eosinophilia, and
eosinophil-related disorders. Allergy Proc 10: 47-62.
Parsons JC, Bowman DD, Grieve RB 1986. Tissue localization of
excretory-secretory antigens of larval Toxocara canis in acute and
chronic murine toxocariasis. Am J Trop Med Hyg 35: 974-981.
Parsons JC, Coffman RL, Grieve RB 1993. Antibody to interleukin-5
prevents blood and tissue eosinophilia but not liver trapping in murine
larval toxocariasis. Parasite Immunol 15: 501-508.
Rennick DM., Thompson-Snipes L, Coffman RL, Seymour BWP, Jackson JD,
Hudak S 1990. In vivo adminstration of antibody to interleukin-5 inhibits
increased generation of eosinophils and their progenitors in bone marrow
of parasitized mice. Blood 76: 312-316.
Sehmi R, Cromwell O, Wardlaw AJ, Moqbel R, Kay AB 1993. Interleukin-8
is a chemo-attractant for eosinophils purified from subjects with a blood
eosinophilia but not from normal healthy subjects. Clin Exper Allergy:
23: 1027-1036.
Sheron N, Williams R 1992. IL-8 as a circulating cytokine: induction by
recombinant tumor necrosis factor-alpha. Clin Exp Immunol 89:
100-103.
Steel C, Nutman TB 1993. Regulation of Il-5 in onchocerciasis: a
critical role for IL-2. J Immunol 150: 5511-5518.
Yamaguchi Y, Matsui T, Kasahara T 1990. In vivo changes of
hemapoietic progenitors and expression of the interleukin 5 gene in
eosinophilic mice infected with Toxocara canis. Exp Hematol 18:
1152-1157
Warringa RA, Schweizer RC, Maioke T, Kuijper PH, Bruijnzeel,
Koendermann K 1992. Modulation of eosinophil chemotaxis by interleukin-5.
Am J Respir Cell Mol Biol 7: 631-636.
Weg VB, Walsh DT, Faccioli LH, Williams TJ, Feldmann M, Nourshargh S
1995. LPS-induced ^111I-eosinophil accumulation in guinea-pig skin:
evidence for a role for TNFa. Immunology 84: 36-40.
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc97175e.jpg]
[oc97175c.jpg]
[oc97175a.jpg]
[oc97175d.jpg]
[oc97175f.jpg]
[oc97175b.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}