
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 92, Num. s2, 1997, pp. 141-146
|
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92, Suppl. II, pp. 141-146
Modulation by IL-10 of Antigen-induced Allergic Responses in Mice
Claudia Zuany-Amorim^+, B Boris Vargaftig, Marina Pretolani
Unite de Pharmacologie Cellulaire, Institut Pasteur, INSERM n 485, 25, rue
du Dr. Roux, 75015 Paris, France
^+Corresponding author. Fax +33-1-4568.8703. E-mail: czamorim@pasteur.fr
Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97179
Sizes of Files:
Text: 27.7K
Graphics: Line drawings and photographs (jpg) - 117.1K
Over the last few years, we examined the anti-allergic properties of
interleukin (IL)-10 in different models of inflammation in the mouse, as
well as against IgE-dependent activation of mouse bone marrow-derived mast
cells (BMMC). We showed that IL-10, concurrently administered with
ovalbumin, inhibited inflammatory cell accumulation in the airways and in
the peritoneal cavity of sensitized mice, as well as the accompanying
cytokine release. IL-10 also blocked antigen-induced cytokine generation by
IgE-stimulated BMMC. Together, these results identify a novel biological
property of IL-10, as a cytokine with potent anti-allergic activities.
Key words: interleukin-10 - eosinophils - mast cells - TNF-alpha -
interleukin-5
Modulation by IL-10 of Antigen-induced Allergic Responses in Mice (Full
Text)
Allergic inflammation involves the attraction and activation of a variety
of cell types at the site of antigen stimulation, including mast cells,
eosinophils, neutrophils and lymphocytes (Arm & Lee 1992). Attention is
particularly focused on eosinophils, since their recruitment in elevated
numbers into the airways of asthmatics after allergen exposure (Bousquet
et al. 1990, Azzawi et al. 1992) may lead to a long-term damage of the
bronchial mucosa (Gleich & Adolphson 1986), as a result of the release of
cytotoxic proteins (Gleich 1990). Several mechanisms responsible for the
attraction and localization of inflammatory cells at site of allergic
reactions have been proposed. Among them, attention is presently focused on
Th2-derived cytokines, particularly interleukin (IL)-5, which promotes the
growth, differentiation, survival and activation of eosinophils (Sanderson
et al. 1985, Clutterbuck et al. 1989, Sanderson 1992) and IL-4, which
induces IgE production by B cells (Pene et al. 1988). In murine models of
allergic inflammation, infiltration of the airways by eosinophils is
regulated by IL-5 and IL-4, since sensitized mice treated with anti-IL-5
(Nakajima et al. 1992) or anti-IL-4 (Lukacs et al. 1994) antibodies display
reduced eosinophilia in their bronchoalveolar lavage (BAL) fluid and
bronchial tissue following antigen challenge.
IL-10 was initially characterized as a product of CD4^+ T-lymphocytes of
the Th2 subtype and shown to inhibit interferon (IFN)-gamma prodution by
Th1 clones (Fiorentino et al. 1989). Several properties of IL-10 on various
cell types have been described since then, including growth stimulation of
thymocytes, mast cells and B cells, and inhibition of cytokine production
by activated monocytes, macrophages (Moore et al. 1993) and, more recently,
by neutrophils (Cassatella et al. 1993, Kasama et al. 1994) and eosinophils
(Takanashi et al. 1994). Only a few studies, however, have focused on the
in vivo allergic modulatory properties of IL-10.
In this report, we evaluated the effects of IL-10 on: (i) Antigen-induced
airway inflammation in immunized BALB/c mice; (ii) IgE-dependent activation
of mouse bone marrow-derived mast cells (BMMC); (iii) Antigen-induced IL-5
generation, CD4^+ T-lymphocyte infiltration and activation into the mice
peritoneal cavity.
Antigen-induced cellular recruitment into the airways of sensitized mice
- The intra-nasal instillation of 10 ug ovalbumin to sensitized BALB/c
mice induced a rise in the number of eosinophils in the bronchial tissue at
24 hr (Fig. 1A). Treatment with 0.1 ug recombinant murine (rm) IL-10,
markedly decreased ovalbumin-induced eosinophil infiltration in the
peribronchial wall (Fig. 1B), as well as in the BAL fluid (Zuany-Amorim et
al. 1995).
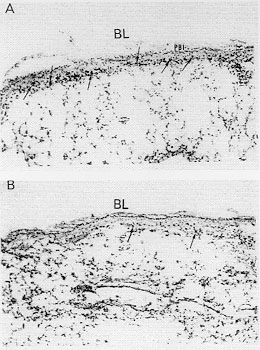
Figure 1:
cyanide-resistant eosinophil peroxidase
activity in the bronchial wall of sensitized saline- or antigen-challenged
mice. (A) Lung section from an immunized ovalbumin-challenged mouse used 24
hr after the challenge, showing massive eosinophil infiltration
concentrated in the bronchial submucosa (arrows); (B)
ovalbumin-challenged-rmIL-10 (0.1 ug/mouse)-treated mouse with scant
eosinophils in the bronchial submucosa (arrows). BL : bronchial lumen, PBI
: peribronchial infiltrate. Original magnifications x 100.
Several lines of evidence designate tumor-necrosis factor (TNF)-alpha as a
cytokine generated by numerous cell types during inflammatory reactions and
shock states (Tracey & Cerami 1993). The participation of TNF-alpha in
allergic reactions is linked to different observations, including its
release by mast cells, basophils and alveolar macrophages upon
IgE-dependent mechanisms (Burd et al. 1989, Gordon & Galli 1990, Ohno et
al. 1990, Gosset et al. 1991). High levels of TNF-alpha have been
shown in monocytes and epithelial cells from asthmatics (Mattoli et al.
1991) and BAL fluid from antigen-challenged guinea-pigs (Watson et al.
1993). Finally, treatment of sensitized guinea-pigs with an IL-1 receptor
antagonist prevents antigen-induced eosinophil accumulation and TNF-alpha
generation in the BAL fluid (Watson et al. 1993), indicating that TNF-alpha
may modulate allergic airway inflammation. We thus investigated whether
antigen challenge induced the release of this cytokine in the BAL fluid of
sensitized mice and verified the potential modulatory activity of IL-10 in
this process. The intra-nasal administration of ovalbumin was followed by a
marked rise in the levels of TNF-alpha in the BAL fluid, which reached a
peak at 1 hr to resolve between 3 and 6 hr after the challenge (Fig. 2A).
The simultaneous administration of ovalbumin and rmIL-10 indicated a
substantial reduction in the levels of TNF-alpha, particularly at 1 hr (
Fig. 2A), a result extending previous observations showing reduction by
IL-10 of LPS-induced in vivo TNF-alpha generation (Howard et al.
1993, Takanashi et al. 1994). Thus, the observation that TNF-alpha
released upon antigenic stimulation is also susceptible of inhibition by
IL-10 may have important consequences for the subsequent invasion of the
bronchial wall by inflammatory cells.
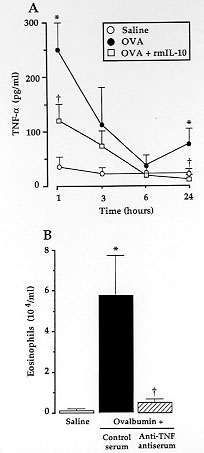
Figure 2 - A:
inhibition by rmIL-10 of antigen-induced
TNF-alpha generation in the BAL fluid from sensitized mice. Sensitized
BALB/c mice were challenged with intra-nasal saline (O), or with 10 ug
ovalbumin, administered either alone (solid circle) or with 0.1 (open
square) ug rmIL-10 and they were killed at various time-intervals after the
challenge. The levels of TNF-alpha in the BAL fluid were evaluated by a
double-sandwitch specific ELISA. Results are expressed as means +/- SEM of
5-9 experiments for each group. B: eosinophil distribution in the BAL fluid
from sensitized mice challenged with intra-nasal saline (open square) or
ovalbumin and treated concomitantly, via the intra-nasal route, with
50 ul/mouse of a sheep anti-murine TNF-alpha antiserum (grey square) or
with its control preimmune serum (solid square). Results represent the
number of eosinophils x 10^4/ ml BAL fluid. * p < 0.05 and ^+
p < 0.05, as compared to saline- or ovalbumin-challenged untreated
mice, respectively.
Accordingly, failure by IL-10 to modify antigen-induced cellular
infiltration when administered 1 hr after the challenge, i.e., the time of
the peak release of TNF-alpha in the BAL fluid, support the hypothesis that
blockade of local TNF-alpha generation and inhibition of leukocyte
recruitment are related phenomena. In confirmation, we demonstrate that the
local administration of an antiserum to TNF-alpha markedly reduced
antigen-induced eosinophil accumulation in the BAL fluid ( Fig. 2B). These
results extend those from previous studies showing the participation of
TNF-alpha in leukocyte infiltration associated with IgE-dependent cutaneous
inflammation in sensitized mice (Wershil et al. 1991). Together, these
results suggest that the inhibitory effect of IL-10 probably involved
down-regulation of TNF-alpha generation in the BAL fluid, since treatment
of sensitized mice with a specific anti-TNF-alpha antiserum drastically
reduced airway eosinophilia.
IgE-dependent cytokine generation by mast cells - Contrary to the
inhibition of allergic airway inflammation observed when rmIL-10 was given
by intra-nasal route, its subcutaneous injection did not modify
antigen-induced cellular accumulation in the BAL fluid (data not shown).
These results suggest that the target(s) for IL-10 are located in the
airways and that the amounts of rmIL-10 reaching the bronchial compartment
after its systemic administration are probably not sufficient to display an
effect. Since mast cells are one of the major sources of TNF-alpha in the
bronchial wall, we hypothesized that reduced TNF-alpha release observed in
the BAL fluid from antigen challenged IL-10-treated mice resulted from mast
cell deactivation by IL-10. To address this question, we tested the extent
to which IL-10 could interfere with IgE-mediated in vitro mast cell
activation, with particular emphasis to cytokine production.
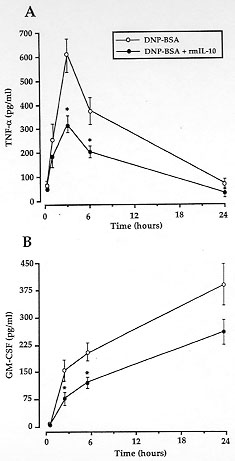
Figure 3: effect of
rmIL-10 on DNP-BSA-induced TNF-
alpha (A) and GM-CSF (B) release from mouse BMMC. IgE-sensitized BMMC were
incubated with 50 ng/ml rmIL-10 (solid circle) for 10 min and then
challenged with 10 ng/10^6 cells of DNP-BSA (O). The levels of TNF-alpha
and GM-CSF were determined by a specific ELISA. Values are means +/- SEM of
4-8 experiments. *^ p < 0.05, as compared with sensitized
DNP-BSA-stimulated cells.
Dinitrophenyl-bovine serum albumin (DNP-BSA)-mediated stimulation of BMMC
was followed by TNF-alpha release, which occurred as early as 1 hr after
challenge, peaked at 3 hr and resolved between 6 and 24 hr (Fig. 3A).
Incubation of the cells with 50 ng/ml rmIL-10 resulted in a significant
inhibition of TNF-alpha production, particularly at 3 and 6 hr (Fig. 3A).
These findings suggest that TNF-alpha originating from pulmonary mast cells
upon antigenic stimulation is one of the main targets for the anti-allergic
activity of IL-10. Northern blot analysis revealed a reduction by rmIL-10
of TNF-alpha mRNA expression, a finding consistent with the inhibition of
TNF-alpha protein release (Arock et al. 1995). In contrast, while GM-CSF
protein release in the medium was decreased by more than 50 % by 50 ng/ml
of rmIL-10 (Fig. 3B), the same concentration of the cytokine failed to
reduce GM-CSF mRNA expression (Arock et al. 1995). This suggests that IL-10
interferes rather with post-transcriptional events involved in GM-CSF
production than with its gene expression.
Antigen-induced IL-5 generation, CD4^+ T-lymphocyte and
eosinophil infiltration into the mice peritoneal cavity - The i.p.
injection of 1 ug ovalbumin to sensitized BALB/c mice induced a marked
increase in the number of eosinophils, starting at 6 hr and reaching a
plateau between 24 and 48 hr (Fig. 4). No changes in the number of
eosinophils were observed in saline-challenged mice at any time point (
Fig. 4A).
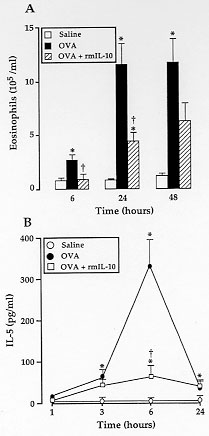
Figure 4 - A: effect
of rmIL-10 on antigen-induced
eosinophil accumulation in the PL fluid from sensitized mice killed 6, 24
or 48 hr after the challenge. Animals were injected i.p. either with saline
(open square), or with 1 ug ovalbumin alone (solid square), or mixed with
0.1 ug/cavity rmIL-10 (grey square). B: inhibition by rmIL-10 of
antigen-induced IL-5 release in the PL fluid from sensitized mice.
Sensitized mice were challenged with saline (O), or with 1 ug ovalbumin
alone (solid circle), or concurrently administered with 0.1 ug rmIL-10
(open square) and they were sacrificed at various time intervals after the
challenge. The levels of IL-5 in the PL fluid were determined by enzyme
immunometric assay. Results are expressed as means +/- SEM (vertical bars)
of 3-14 experiments.
*p < 0.05 and + p < 0.05, as
compared to saline- or ovalbumin-challenged untreated mice, respectively.
The administration of 0.1 ug rmIL-10, concomitantly injected with 1 ug
ovalbumin, reduced eosinophil counts at 6 h and 24 hr after the antigen
challenge. At 48 hr, even though the numbers of eosinophils were reduced by
47% by the treatment with rmIL-10, the high variability in cell counts
precluded the results from achieving statistical significance (Fig. 4A).
Contrary to what we have observed when the animals were challenged
via the intra-nasal route, no release of TNF-alpha was detected in
the supernatant of peritoneal lavage (PL) fluid after the intra-peritoneal
administration of ovalbumin, at any time-point (data not shown). These
results indicate that IL-10 reduced antigen-induced eosinophilia in the
peritoneal cavity by mechanisms unrelated to the blockade of the local
generation of TNF-alpha and suggest that, depending upon the site of
inflammation, IL-10 may exhibit anti-allergic properties by acting on
different cell types. Interestingly enough, the administration of rmIL-10,
3 hr after antigen challenge still inhibits eosinophilia in the PL fluid at
24 hr (Zuany-Amorim et al. 1996). This result differs from our previous
findings showing that the intra-nasal instillation of rmIL-10, 1 hr after
ovalbumin stimulation, failed to protect sensitized mice against eosinophil
accumulation in the BAL fluid (Zuany-Amorim et al. 1995). This discrepancy
suggests that mediator(s) and/or cytokine(s) released in the peritoneal
cavity later than 3 hr after ovalbumin stimulation are involved in the
eosinophil recruitment and are target for the anti-allergic activity of
IL-10. The data presented herein show marked down-regulation by rmIL-10 of
antigen-induced IL-5 release in the PL fluid at 6 hr (Fig.
4B), a phenomenon temporally correlated with the subsequent inhibition
of eosinophil accumulation in this compartment. The IL-5-dependency of
allergic eosinophilia in sensitized mice has been widely demonstrated
(Kaneko et al. 1991, Okudaira et al. 1991). In particular, the observation
that the in vivo administration of an antibody to murine IL-5
receptor prevented eosinophilia in IL-5 transgenic mice (Hitoshi et al.
1990), further supports the concept that IL-5 generation and eosinophil
recruitment are related events. In confirmation of these findings, we
showed a reduced antigen-induced eosinophil accumulation in the peritoneal
cavity of sensitized mice after treatment with a neutralizing anti-IL-5
monoclonal antibody (Zuany-Amorim et al. 1996).
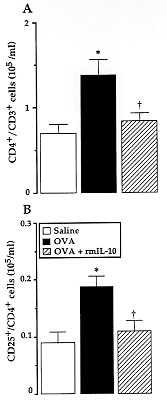
Figure 5: effect of
rmIL-10 on antigen-induced
CD4^+/CD3^+ and CD25^+/CD4^+ T cell accumulation in the PL fluid from
sensitized mice killed 24 hR after the challenge. Animals were injected
i.p. either with saline (open square), or with 1 ug ovalbumin alone (solid
square), or mixed with 0.1 ug/cavity rmIL-10 (grey square). Results are
expressed as means +/- SEM (vertical bars) of 5-11 experiments.
*p < 0.05 and + p < 0.05, as compared to saline- or
ovalbumin-challenged untreated mice, respectively.
Ovalbumin stimulation was followed by a time-dependent rise in CD4^+ T
cells at 24 hr (Fig. 5A). In parallel, a significant increment in the
number of CD4^+ cells bearing IL-2 receptor on their surface (CD25^+/CD4^+
T-cells) was observed at 24 hr (Fig. 5B). When 0.1 ug rmIL-10 were
co-injected with ovalbumin, a significant reduction in the number of CD4^+
T-lymphocytes and in that of CD25^+/CD4^+ cells at 24 hr was observed (
Fig. 5A, B). These results parallel those of Jinquan et al. (1993), who
demonstrated that IL-10 inhibits in vitro CD4^+ T-lymphocyte
chemotaxis induced by IL-8.
Accumulating evidence indicates that the recruitment and function of
eosinophils may be orchestrated by the products of activated T-lymphocytes.
Thus, our observations that IL-5 release in the PL fluid rised at 6 hr,
while T-cell numbers increased at 24 hr, suggest that T-lymphocytes
residing in the peritoneal cavity, rather than those having infiltrated
this compartment in response to antigen challenge, are responsible for the
early cytokine release and the accompanying eosinophilia. Alternatively,
cell types other than T-lymphocytes, such as mast cells (Plaut et al.
1989), may be involved in ovalbumin-induced IL-5 production at 6 hr.
Nevertheless, our observation that specific depletion of CD4^+
T-lymphocytes abolished both eosinophil accumulation and IL-5 release in
the peritoneal cavity (Zuany-Amorim et al. 1996) suggests that IL-5
originating from T-cells plays an important role in antigen-induced
eosinophilia in this model.
To verify whether IL-10 would directly interfere with T-cell activation, we
triggered in vitro and in vivo the CD3/Tcell receptor complex
with an anti-CD3 mAb (Hirsch et al. 1989). In vivo stimulation of
non immunized mice with anti-CD3 mAb led to a time-dependent production of
IL-5 in the PL fluid, which was not modified by treatment of the animals
with a dose of rmIL-10 effective against ovalbumin-induced eosinophilia and
IL-5 release. In parallel, IL-10 failed to inhibit CD3-dependent IL-5
generation from purified spleen CD4^+ T-lymphocytes (Zuany-Amorim et al.
1996). These findings suggest that mechanisms unrelated to T-cell
activation (possibly antigen-presenting cell deactivation) are targets for
the anti-allergic activity of IL-10 in this model.
In recent years, evidence has suggested that allergic inflammation is a
Th2-cytokine-mediated disease. As IL-10 is produced by bronchial epithelial
cells (Bonfield et al. 1995) and can suppress acute inflammation induced by
immune-complexes in mice lungs (Shanley et al. 1995), there has been
considerable interest in the role of IL-10 in regulating Th2-like responses
leading to allergic lung inflammation. Our present results showing that
IL-10 can negatively regulate allergic inflammatory reaction by inhibiting
IL-5 and TNF-alpha release, suggest that this cytokine may have therapeutic
value for treating eosinophilic disorders, such as bronchial asthma.
REFERENCES
Arm JP, Lee TH 1992. The pathobiology of bronchial asthma. Adv
Immunol 51: 323-382.
Arock M, Zuany-Amorim C, Singer M, Benhamou M, Pretolani M 1995.
Interleukin-10 inhibits mast cell activation. Eur J Immunol 26:
166-172.
Azzawi M, Johnston S, Majumdar S, Kay AB, Jeffery PK 1992. T
lymphocytes and activated eosinophils in airway mucosa in fatal asthma and
cystic fibrosis. Am Rev Respir Dis 145: 1477-1486.
Bonfield TL, Konstan MW, Burfeind JR, Panuska JB, Hilliard JB, Berger M
1995. Normal bronchial epithelial cells constitutively produce the
anti-inflammatory cytokine interleukin-10, which is downregulated in
cystic fibrosis. Am J Respir Cell Mol Biol 13: 257-261.
Bousquet J, Chanez P, Lacoste JY, Barneron G, Ghavanian N, Enander I,
Venge P, Ahlstedt S, Simony LJ, Godard P, Michel FB 1990. Eosinophilic
inflammation in asthma. N Engl J Med 323: 1033-1039.
Burd PR, Rogers HW, Gordon JR, Martin CA, Jayaraman S, Wilson SD,
Dvorak AM, Galli SJ, Dorf ME 1989. Interleukin-3- dependent and
-independent mast cells stimulated with IgE and antigen express multiple
cytokines. J Exp Med 170: 245-257.
Cassatella MA, Meda L, Bonora S, Ceska M, Constantin G 1993.
Interleukin-10 (IL-10) inhibits the release of proinflammatory cytokines
from human polymorphonuclear leukocytes. Evidence for an autocrine role of
tumor necrosis factor and IL-1b in mediating the production of IL-8
triggered by lipopolysaccharide. J Exp Med 178: 2207-2211.
Clutterbuck EJ, Hirst EM, Sanderson CJ 1989. Human interleukin-5 (IL-5)
regulates the production of eosinophils in human bone marrow cultures:
comparison and interaction with IL-1, IL-3, IL-6, and GM-CSF. Blood
73: 1504-1512.
Fiorentino DF, Bond MW, Mosmann TR 1989. Two types of mouse T helper
cells: IV. Th2 clones secrete a factor that inhibits cytokine production
by Th1 clones. J Exp Med 170: 2081-2095.
Gleich GJ, Adolphson CR 1986. The eosinophilic leukocyte: Structure and
function. Adv Immunol 39: 177-183.
Gleich GJ 1990. The eosinophil and bronchial asthma: current
understanding. J Allergy Clin Immunol 85: 422-427.
Gordon JR, Galli SJ 1990. Mast cells as a source of both preformed and
immunologically inducible TNF-alpha/cachectin. Nature 346:
274-276.
Gosset P, Tsicopoulos M, Wallaert B, Vannimenus C, Joseph M, Tonnel AB,
Capron A 1991. Increased secretion of tumor necrosis factor a and
interleukin-6 from alveolar macrophages consecutive to the development of
the late asthmatic reaction. J Allergy Clin Immunol 88: 561-571.
Hirsch R, Gress RE, Pluznik DH, Eckhaus M, Bluestone JA 1989. Effects
of in vivo administration of anti-CD3 monoclonal antibody on T cell
function in mice. II. In vivo activation of T cells. J
Immunol 142: 737-743.
Hitoshi Y, Yamaguchi N, Korenaga M, Mita S, Tominaga A, Takatsu K 1990.
In vivo administration of antibody to murine IL-5 receptor inhibits
eosinophilia of IL-5 transgenic mice. Int Immunol 3: 135-139.
Howard M, Muchamuel T, Andrade S, Menon S 1993. Interleukin 10 protects
mice from lethal endotoxemia. J Exp Med 177: 1205-1208.
Jinquan T, Larsen CG, Gesser B, Matsushima K, Thestrup-Pedersen K 1993.
Human IL-10 is a chemoattractant for CD8^+ T lymphocyte and an inhibitor
of IL-8-induced CD4^+ T lymphocyte migration. J Immunol 151:
4545-4551.
Kaneko M, Hitoshi Y, Takatsu K, Matsumoto S 1991. Role of interleukin-5
in local accumulation of eosinophils in mouse allergic peritonitis. Int
Arch Allergy Appl Immunol 96: 41-45.
Kasama T, Strieter RM, Lukacs NW, Burdick MD, Kunkel SL 1994.
Regulation of neutrophil-derived chemokine expression by IL-10. J
Immunol 152: 3559-3569.
Lukacs WN, Strieter RM, Chensue SW, Kunkel SL 1994.
Interleukin-4-dependent pulmonary eosinophil infiltration in a murine
model of asthma. Am J Respir Cell Mol Biol 10: 526-532.
Mattoli S, Mattoso VL, Soloperto M, Allegra L, Fasoli A 1991. Cellular
and biochemical characteristics of bronchoalveolar lavage fluid in
symptomatic nonallergic asthma. J Allergy Clin Immunol 87: 794-802.
Moore KW, O'Garra A, de Waal Malefyt R, Vieira P, Mosmann TR 1993.
Interleukin-10. Annu Rev Immunol 11: 165-190.
Nakajima H, Iwamoto I, Tomoe S, Matsumura R, Tomioka H, Takatsu K,
Yoshida S 1992. CD4^+ T-lymphocytes and interleukin-5 mediate
antigen-induced eosinophil infiltration into the mouse trachea. Am Rev
Respir Dis 146: 374-377.
Ohno I, Yamaguchi K, Takasihima T 1990. Gene expression and production
of tumor necrosis factor by a rat basophilic cell line (RBL-2H3) with IgE
receptor triggering. Immunology 70: 88-93.
Okudaira H, Matsuzaki M, Dohl G, Suko M, Kasuya M, Takatsu K 1991.
T-cell-dependent accumulation of eosinophils in the lung and its
inhibition by monoclonal anti-interleukin-5. Int Arch Allergy Appl
Immunol 199: 171-173.
Pene J, Rousset F, Brière F, Chretien I, Bonnefoy JY, Spits HT, Yokota
N, Arai KI, Banchereau J, de Vries JE 1988. IgE production by human B
cells is induced by IL-4 and suppressed by interferons g and a and
prostaglandin E2. Proc Natl Acad Sci USA 85: 6880-6884.
Plaut M, Pierce JH, Watson CJ, Hamley-Hyde J, Nordan RP, Paul WE 1989.
Mast cell lines produce lymphokines in response to cross-linkage of FceRI
or to calcium ionophores. Nature 339: 64-67
Sanderson CJ, Warren DJ, Strath M 1985. Identification of a lymphokine
that stimulates eosinophil differentiation in vitro. Its
relationship to IL-3 and functional properties of eosinophils produced in
cultures. J Exp Med 162: 60-74.
Sanderson CJ 1992. Interleukin-5, eosinophils, and disease.
Blood 79: 3101-3109.
Shanley TP, Schmal H, Friedl HP, Ward P 1995. Regulatory effects of
intrinsic IL-10 in IgG in immune complex-induced lung injury. J
Immunol 154: 3454-3460.
Takanashi S, Nonaka R, Xing Z, O'Byrne P, Dolovich J, Jordana M 1994.
Interleukin 10 inhibits lipopolysaccharide-induced survival and cytokine
production by human peripheral blood eosinophils. J Exp Med 180:
711-715.
Tracey KJ, Cerami A 1993. Tumor necrosis factor, other cytokines and
disease. Annu Rev Cell Biol 9: 317-343.
Watson ML, Smith D, Bourne AD, Thompson RC, Westwick J 1993. Cytokines
contribute to airway dysfunction in antigen-challenged guinea-pigs:
inhibition of airway hyperreactivity, pulmonary eosinophil accumulation,
and tumor necrosis factor generation by pretreatment with an interleukin-1
receptor antagonist. Am J Respir Cell Mol Biol 8: 365-369.
Wershil BK, Wang Z, Gordon JR, Galli SJ 1991. Recruitment of
neutrophils during IgE-dependent cutaneous late phase responses in the
mouse is mast cell dependent: partial inhibition of the reaction with
antiserum against tumor necrosis factor-alpha. J Clin Invest
87: 446-453.
Zuany-Amorim C, Haile S, Leduc D, Dumarey C, Huerre M, Vargaftig BB,
Pretolani M 1995. Interleukin-10 inhibits antigen-induced cellular
recruitment into the airways of sensitized mice. J Clin Invest
95: 2644-2650.
Zuany-Amorim C, Creminon C, Nevers MC, Nahori MA, Vargaftig BB,
Pretolani M 1996. Modulation by IL-10 of antigen-induced IL-5 generation,
CD4^+ T-lymphocyte and eosinophil infiltration into the mice peritoneal
cavity. J Immunol 157: 377-384.
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc97179b.jpg]
[oc97179a.jpg]
[oc97179d.jpg]
[oc97179e.jpg]
[oc97179c.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}