
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 92, Num. s2, 1997, pp. 147-155
|
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92, Suppl. II, pp. 147-155
Role of the Cyclosporin-sensitive Transcription Factor NFAT1 in the
Allergic Response
Joao PB Viola, Anjana Rao^+
The Center for Blood Research and the Department of Pathology, Harvard
Medical School, 200 Longwood Ave., Boston, MA 02115, USA
^+Corresponding author. Fax: +617-278.3280. E-mail: arao@cbr.med.harvard.edu
Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97180
Sizes of Files:
Text: 44.6K
Graphics: Line drawings and photographs (jpg) - 60.2K
Proteins belonging to the NFAT (nuclear factor of activated T cells)
family of transcription factors are expressed in most immune cell types,
and play a central role in the transcription of cytokine genes, such as
IL-2, IL-4, IL-5, IL-13, IFN-gamma, TNF-alpha, and GM-CSF. The activity of
NFAT proteins is regulated by the calcium/calmodulin-dependent phosphatase
calcineurin, a target for inhibition by CsA and FK506. Recently, two
different groups have described that mice lacking the NFAT1 transcription
factor show an enhanced immune response, with tendency towards the
development of a late Th2-like response. This review evaluates the possible
role of NFAT proteins in the Th2 immune response and in the
eosinophil-mediated allergic response.
Key words: nuclear factor of activated T cells - interleukin -
cyclosporin
The cytokine profiles of T cells differentiating down the Th1 and Th2
pathways have been described, Th1 cells preferentially produce IL-2 and
IFN-g, whereas Th2 cells produce IL-4, IL-5, IL-10 and IL-13 (Paul & Seder
1994, Carter & Dutton 1996). In many pathological situations, the balance
between Th1 and Th2 immune response determines the outcome of different
immunologically-mediated clinical syndromes including infectious,
autoimmune, and allergic diseases (Carter & Dutton 1996).
Allergic disease is a broad range of disorders including rhinitis,
conjunctivitis, systemic anaphylaxis, and asthma (Casolaro et al. 1996,
Drazen et al. 1996). Atopic allergy is characterized by increased synthesis
of IgE antibodies through the actions of IL-4 and IL-13 in B cell Ig
isotype class switching, directed at groups of antigens that activate the
CD4-dependent Th2-like immune response (Romagnani 1995). The IgE produced
binds to Fce receptors present on the surface of mast cells and basophils,
priming them for activation by antigen, and triggers the release of
vasoactive mediators, chemotactic factors and cytokines (Romagnani 1995,
Drazen et al. 1996). In addition, eosinophils are also involved in the
pathogenesis of allergic reactions, as these cells accumulate at the sites
of allergic inflammation and significantly contribute to the tissue damage
(Desreumaux & Capron 1996).
In asthma, some data suggest that the severity of the disease is related to
the degree of inflammation (Peters 1990, Broide et al. 1991, Pare & Bai
1995), and that the magnitude of the asthmatic response is related to the
number of eosinophils present in the lung (Bradley et al. 1991). Moreover,
suppression of eosinophil accumulation at the site of inflammation impairs
the development of asthma disease (Wagner et al. 1990, Foster et al. 1995).
These data suggest that eosinophils could be a central mediator of the
pathogenesis of allergic disease. We can hypothesize three sequential and
interacting events for how eosinophils mediate inflammation at the site of
allergic response, described as follows: first, eosinophil differentiation
and maturation in the bone marrow; second, rolling, adhesion, and migration
in the inflamed vascular endothelium; and third, activation and
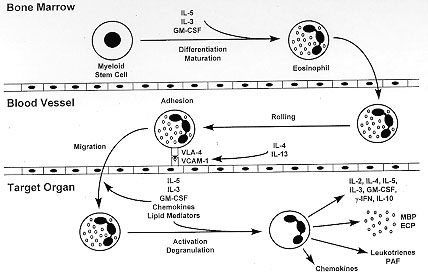
degranulation in the target organ (Fig. 1).
Figure 1: a
simplified view of eosinophil
differentiation and maturation, in the bone marrow; rolling, adhesion and
migration, in the blood vessel; and activation and degranulation, in the
target organ. Cytokines, growth factors, adhesion molecules, chemokines,
and lipid mediators that play a role in each step are indicated.
Eosinophil tissue infiltration is coordinated by an interacting network of
cytokines, chemokines, adhesion molecules, and inflammatory mediators. In
fact, GM-CSF, IL-3 and IL-5 have been described as factors that induce
differentiation, maturation and proliferation of bone marrow eosinophils
(Sanderson et al. 1985, Lopez et al. 1986, Clutterbuck & Sanderson 1988,
Takatsu et al. 1988, Warren & Morre 1988, Yamaguchi et al. 1988). However,
blood eosinophilia is not related to eosinophil tissue accumulation (Dent
et al. 1990, Desreumaux et al. 1996), suggesting that overproduction of
eosinophils is not enough for tissue infiltration by these cells, and
chemoattractant production at the site of inflammation is essential for
eosinophil recruitment.
Rolling and adhesion of eosinophils on vascular endothelium is the first
step for eosinophil infiltration in the target organ, and may depend on
several adhesion molecules (Desreumaux & Capron 1996). However, eosinophils
are the only granulocytes that express VLA-4, and may selectively bind to
endothelial cells via VCAM-1 (Weller et al. 1991, Pretolani et al. 1994,
Nakajima et al. 1994, Wardlaw et al. 1994). Moreover, it has been described
that IL-4 and IL-13 upregulate VCAM-1 on human endothelial cells (Bochner
et al. 1995), suggesting that interaction between VLA-4/VCAM-1 play a
central role in eosinophil migration during the allergic response.
Eosinophil migration into inflamed tissue involves several chemoattractant
mediators, including cytokines, chemokines and lipid mediators, and occurs
after adhesion to the vascular endothelium (Desreumaux & Capron 1996).
GM-CSF, IL-3 and IL-5, are the key cytokines influencing eosinophil
migration and activation (Broide et al. 1992, Weller 1993, Sullivan &
Broide 1996). In fact, several reports demonstrated that IL-5 plays the
central role in eosinophil-mediated allergic responses, since this cytokine
is a selective chemoatractant for eosinophils (Sehmi et al. 1992), and has
the ability to prime and activate these cells (Coeffier et al. 1991, Sehmi
et al. 1992, Warringa et al. 1992). In addition, IL-5 deficient mice do not
show eosinophilia (Kopf et al. 1996), and fail to develop airway
hyperresponsiveness and eosinophil infiltration in an experimental model of
asthma (Foster et al. 1995). Other important chemoattractants and
activators of eosinophils are the C-C subfamily of chemokines (Desreumaux &
Capron 1996). The eosinophil active chemokines include RANTES, MCP-2,
MCP-3, MCP-4, MCP-5, MIP-1a, and eotaxin (Jia et al. 1996, Kita & Gleich
1996, Sarafi et al. 1997). Eotaxin, first described in guinea pigs and
subsequently in mice and humans, is a potent and specific eosinophil
chemoattractant (Jose et al. 1994, Gonzalo et al. 1996, Ponath et al.
1996), and disruption of the eotaxin gene partially reduces tissue
eosinophil infiltration in a model of allergic response (Rothenberg et al.
1997).
Once eosinophils infiltrate the inflamed tissue, they degranulate and
secrete several proinflamatory mediators and cytokines (Weller 1993).
Activated eosinophils release their granule proteins, including the major
basic proteins, eosinophil peroxidase (EPO), eosinophil cationic protein
(ECP), and eosinophil-derived neurotoxin (Desreumaux & Capron 1996). They
also secrete lipid mediators, chemokines, and cytokines, which amplify the
response and generate a feedback loop that perpetuates the allergic
inflammatory response (Drazen et al. 1996). Together, these inflammatory
mediators and cytokines generate tissue damage that could be related with
the clinical symptoms of the different allergic diseases.
NFAT TRANSCRIPTION FACTORS IN IMMUNE RESPONSE
Many of the cytokines that regulate eosinophil function are under the
control of proteins belonging to the NFAT (nuclear factor of activated T
cells) family of transcription factors. These proteins play a key role in
the regulation of cytokine gene transcription during the immune response
(Crabtree & Clipstone 1994, Rao 1994, Jain et al. 1995b). The NFAT family
encodes four distinct classes of proteins: NFAT1 (formerly NFATp), NFAT2
(NFATc), NFAT3 and NFAT4 (NFATx) (Rao et al. 1997). NFAT1, the first
identified member of the family, was cloned from murine (Ar-5) and human
(Jurkat) T cell cDNA libraries (McCaffrey et al. 1993, Luo et al. 1996). A
distinct protein, NFATc (NFAT2), later was also cloned from a Jurkat T cell
cDNA library (Northrop et al. 1994). cDNA clones encoding three other NFAT
proteins: NFAT3, NFAT4 and NFATx (isoform of NFAT4), were isolated from
Jurkat T cell, human peripheral blood (PBL) and human thymus cDNA libraries
(Ho et al. 1995, Masuda et al. 1995).
Despite their name, NFAT proteins are expressed not only in T cells, but
also in other classes of immune and non-immune cells. At the protein level,
NFAT1 and NFAT2 are expressed in peripheral T cells and T cell lines, and
NFAT1 is also expressed in B cells, mast cells, NK cells, monocytes and
macrophages (Ho et al. 1994, Aramburu et al. 1995, Ruff & Leach
1995, Wang et al. 1995, Weiss et al. 1996). Moreover, NFAT1 is expressed in
a neuronal cell line and in the nervous system (Ho et al. 1994), and an
endothelial cell line (Cockerill et al. 1995a, Wang et al. 1995). NFAT1 and
NFAT2 mRNAs are expressed in peripheral lymphoid tissue (spleen and PBL),
and NFAT2 mRNA is upregulated in activated T cells and NK cells (Northrop
et al. 1994, Aramburu et al. 1995, Hoey et al. 1995, Masuda et al. 1995,
Park et al. 1996). NFAT4 mRNA is expressed at high levels in the thymus
(Hoey et al. 1995, Ho et al. 1995, Masuda et al. 1995), and NFAT3 is
expressed at low levels in lymphoid tissues (Hoey et al. 1995).
Several isoforms have been described for NFAT1, NFAT2 and NFAT4. Sequence
homology represented in all the isoforms suggests two different domains,
comprising the DNA-binding domain (DBD) and the NFAT homology region (NHR)
(Jain et al. 1995a, Luo et al. 1996). The DBD, which is located between
amino acid residues 400 and 700, is highly conserved within the NFAT
family, and shows moderate sequence similarity to the DNA-binding domains
of Rel-family proteins (Nolan 1994, Jain et al. 1995a, Chytil & Verdine
1996). This domain contains the highly conserved RAHYETEG sequence in which
residues contact DNA (Jain et al. 1995a, Chytil & Verdine 1996). The NHR is
located in the N-terminal region, comprising 300 amino acids, and shows a
strong conservation of several sequence motifs characteristic of the NFAT
family (Ho et al. 1995, Hoey et al. 1995, Masuda et al. 1995, Luo et al.
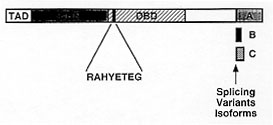
1996) (Fig. 2).
Figure 2: schematic
diagram of the primary structure of
the NFAT1 protein, as deduced from analysis of cDNA clones. The region of
highest homology within NFAT proteins is the DNA-binding domain (DBD),
which shows similarity to the Rel homology region of Rel-family
transcription factors, and encodes the amino acids that contact DNA. Other
regions such as transactivation domain (TAD), NFAT homology region (NHR),
and splicing variants isoforms are indicated.
NFAT transcription factors are cytosolic proteins constitutively expressed
in resting cells (Rao et al. 1997). NFAT proteins are activated by
stimulation of receptors coupled to calcium mobilization, such as the
antigen receptors on T and B cells (Yaseen et al. 1993, Choi et al. 1994,
Crabtree & Clipstone 1994, Rao, 1994, Venkataraman et al. 1994, Jain et al.
1995b, Serfling et al. 1995, Loh et al. 1996b), Fce receptors on mast cells
and basophils (Hutchinson & McCloskey 1995, Prieschl et al. 1995a, Weiss et
al. 1996), the Fcg receptors on macrophages and NK cells (Aramburu et al.
1995), and receptors coupled to heterotrimeric G proteins (Desai et al.
1990, Wu et al. 1995, Boss et al. 1996). Three different steps of
activation have been defined for NFAT proteins: dephosphorylation, nuclear
translocation, and DNA binding. In resting cells, NFAT proteins are
phosphorylated and cytoplasmic, and show a low affinity for DNA (Shaw et
al. 1995). Stimuli that trigger calcium mobilization result in rapid
dephosphorylation of NFAT proteins and their translocation to the nucleus,
and dephosphorylated proteins show increased affinity for DNA (Shaw et al.
1995, Loh et al. 1996a,b). Receptor stimulation and calcium
mobilization result in activation of the calmodulin-dependent phosphatase
calcineurin (Weiss & Littman 1994). Each step of NFAT activation is blocked
by the calcineurin inhibitors CsA or FK506, suggesting that calcineurin is
a major upstream regulator of NFAT proteins, and that dephosphorylation is
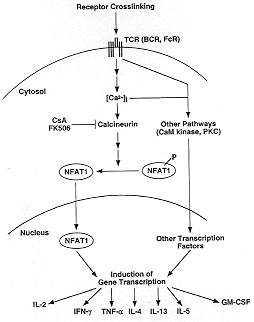
the initial step of NFAT activation (Fig. 3).
Figure 3: signal
transduction mechanisms leading to
transcription of cytokine genes in activated T cells (and other cells of
the immune system) upon stimulation through surface receptors capable of
mobilizing calcium. Abbreviations: TCR, T-cell receptor; BCR, B-cell
receptor; FcR, Fcg and Fce receptors; CsA, cyclosporin A; PKC, protein
kinase C; CaM kinase, calmodulin-dependent kinase; P, phosphorylation.
Stimulated cells inducibly transcribe a large number of genes, such as
genes encoding transcription factors, signalling proteins, cytokines, cell
surface receptors, and other effector proteins (Leonard et al. 1987,
Crabtree 1989, Cockerill et al. 1995b, Kelly & Siebenlist 1995). NFAT was
first identified in T cells as a rapidly-inducible nuclear factor binding
to the distal antigen receptor response element of the human IL-2 promoter
(Shaw et al. 1988). Over the next few years, studies from several
laboratories indicated that the promoter/enhancer regions of different
activation genes possessed binding sites for NFAT family proteins,
including the cytokines IL-2 (Shaw et al. 1988, Mattila et al. 1990, Randak
et al. 1990, Brabletz et al. 1991, Ullman et al. 1991, Jain et al. 1993a,
Rooney et al. 1995a), IL-4 (Casalaro et al. 1995, Rooney et al. 1995b),
IL-5 (Prieschl et al. 1995a,b), IL-13 (Dolganov et al. 1996), IFN-g (Brown
et al. 1991, Campbell et al. 1996), TNF-a (McCaffrey et al. 1994, Tsai et
al. 1996), and GM-CSF(Cockerill et al. 1995a,b), as well as the cell
surface receptors CD40L (Schubert et al. 1995, Tsytsykova et al. 1996), and
CTLA-4 (Perkins et al. 1996). Based on comparison of these sequences, the
NFAT binding site is presented as a 9 bp element, possessing the consensus
sequence (A/T)GGAAA(A/N)(A/T/C)N (Rao et al. 1997). In addition, NFAT
proteins show a characteristic ability to cooperate with AP-1 proteins in
DNA-binding and transactivation (Rao 1994). The interaction between NFAT
proteins and AP-1 involves binding of these transcription factors to
adjacent sites on DNA, and results in stabilisation of the NFAT-DNA
interaction (Jain et al. 1993a,b, Chen et al. 1995). Note that cytokines
that are inhibited by CsA or FK506 have present in their promoter regions
binding sites for NFAT proteins, suggesting that NFAT transcription factors
are major targets of these immunosuppressive drugs (Rao et al. 1997).
Cytokine expression by different cells of the immune system plays a central
role in the immune response, and each cell type produces a characteristic
pattern of cytokines (Paul & Seder 1994). The immune response is
coordinated by an interacting network of transcription factors that dictate
expression of different cytokines (Crabtree 1989, Paul & Seder 1994). To
study the unique functions of the NFAT1 transcription factor in the in
vivo immune response, two different groups generated mutant mice
carrying a disrupted NFAT1 gene (Hodge et al. 1996, Xanthoudakis et al.
1996). Although all reports indicate that the NFAT1 transcription factor is
a positive regulator of cytokine expression, surprisingly, certain primary
and secondary immune responses in mice lacking NFAT1 gene were enhanced,
such as increased intrapleural accumulation of eosinophils and increased
serum IgE levels in an in vivo model of allergic inflammation
(Xanthoudakis et al. 1996), increased serum IgE levels in response to
immunization with TNP-ovalbumin (Hodge et al. 1996), and more efficient
differentiation towards a Th2 phenotype in spleen cells stimulated in
vitro with IL-4 and anti-CD3 (Hodge et al. 1996). In the next section
we discuss the possible role of the NFAT1 transcription factor in the Th2
immune response and eosinophil-mediated allergic inflammation.
ROLE OF NFAT1 TRANSCRIPTION FACTOR IN THE ALLERGIC RESPONSE
The response of the immune system to antigen is coordinated by an
interacting network of transcription factors that dictate expression of
different effector proteins that regulate the immune response (Crabtree
1989, Paul & Seder 1994). However, it is not known how the same stimuli can
be responsible for encoding the specificity of cellular response. Recently,
it has been described that different calcium signalling patterns can
activate different transcription factors, demonstrating that the same
second messenger can drive specificity in signalling to the nucleus
(Dolmetsch et al. 1997). Nevertheless, the molecular basis for the
tissue-specific expression of Th1/Th2-like cytokines has remained elusive.
Over the next few years, several groups have been described important
advances in signaling and gene transcription in the immune system using
in vivo gene disruption.
In other to address the specialized functions of NFAT1 transcription factor
in the in vivo immune response, mutant mice carrying a disrupted
NFAT1 gene have been described (Hodge et al. 1996, Xanthoudakis et al.
1996). In both cases the targeted exon was in the DNA-binding domain
encoding the Rel-homology region (see above), and the disruption resulted
either in the expression of a truncated protein without DNA-binding
activity (Hodge et al. 1996), or in no protein expression (null phenotype)
(Xanthoudakis et al. 1996). Except for a moderate degree of splenomegaly,
NFAT1-deficient mice developed normally, did not exhibit any obvious
behavioral deficiencies, and were immunocompetent.
In the primary immune response, NFAT1-deficient mice showed no impairment
in IL-2, IL-4, IFN-g and TNF-a production by in vitro stimulation of
spleen cells with anti-CD3 antibody or Con A (Xanthoudakis et al. 1996).
However, in an in vivo model of primary response NFAT1-deficient
mice showed an early impairment of several cytokines, such as IL-4, IL-13,
TNF-a and GM-CSF, and cell surface receptors, including CD40L and FasL
(Hodge et al. 1996). These results suggest that the NFAT1 protein played an
important role in the primary in vivo immune response that could not
have been predicted from the in vitro experiments.
Surprisingly, certain primary and secondary immune responses were markedly
enhanced. In fact, CD4 T cells hyperproliferated in an in vitro
response to anti-CD3 antibody, and an in vitro model of T helper
(Th) differentiation, NFAT1-deficient mice showed an increased level of
IL-4 production at later timepoints (Hodge et al. 1996). In addition, these
mice presented high serum IgE levels in response to immunization with
ovalbumin (Hodge et al. 1996, Xanthoudakis et al. 1996). Moreover,
NFAT1-deficient mice consistently showed a marked increase in the secondary
immune response using two different experimental models. First, cells from
draining lymph nodes of mice that had been sensitized with ovalbumin
hyperproliferated after a secondary in vitro stimulation with the
same antigen (Xanthoudakis et al. 1996). Second, an allergic/inflammatory
response to antigen was assessed in vivo. Mice that had been
previously sensitized to ovalbumin were restimulated by intrapleural
injection of antigen, and the accumulation of eosinophils in the pleural
cavity was assessed. NFAT-deficient mice showed a marked increased in the
number of eosinophils in the pleural cavity and a corresponding increase in
the level of serum IgE (Xanthoudakis et al. 1996).
The immune phenotype of NFAT1-deficient mice illustrates three important
points. First, these mice are immunocompetent rather than immunodeficient
and do not show any gross impairment in the production of NFAT-dependent
cytokines, indicating that the lack of NFAT1 is compensated for by the
presence of other NFAT proteins. Second, the increased secondary immune
responses and increased cell proliferation observed in NFAT1-deficient mice
suggests that NFAT1 may actually have an overall negative effect on immune
responsiveness in normal mice. This behaviour is not unprecedented: for
example, in signal transduction pathways, kinases that are activated early
during a response often activate feedback processes that contribute to the
late downregulation of the same response. Finally, the unusual
hyper-eosinophila of NFAT1-deficient mice in a model of allergy, and their
tendency towards the late production of Th2-type cytokines, suggests that
NFAT1 critically influences Th differentiation during the normal immune
response. NFAT1 could act to promote the transcription of genes encoding
immunosuppressive cytokines, cytokines that skew T cell differentiation
towards the Th1 pathway, or cytokines that suppress differentiation towards
the Th2 pathway. Alternatively, NFAT1 could inhibit the production of
cytokines having the opposite effect. These possibilities are not mutually
exclusive. Given the importance of Th1-Th2 cytokine production in asthma,
allergy, and other clinical situations, it is of considerable interest to
understand the mechanisms by which NFAT1 exerts its profound effects on T
cell differentiation and function.nbsp;
ACKNOWLEDGMENTS
To Drs Patricia Bozza and Heidi Okamura for critical reading of this
manuscript. Work in the A Rao laboratory was supported by grants from
National Institute of Health and Hoffman-La Roche, Inc.
REFERENCES
Aramburu J, Azzoni L, Rao A, Perussia B 1995. Activation and expression
of the nuclear factor of activated T cells, NFATp and NFATc, in human
natural killer cells: regulation upon CD16 ligand binding. J Exp
Med 182: 801-810.
Bochner BS, Klunk DA, Sterbinsky SA, Coffman RL, Schleimer RP 1995.
IL-13 selectively induces vascular cell adhesion molecule-1 expression in
human endothelial cells. J Immunol 154: 799-803.
Boss V, Talpade DJ, Murphy TJ 1996. Induction of NFAT-mediated
transcription by Gq-coupled receptors in lymphoid and non-lymphoid
cells. J Biol Chem 271: 10429-10432.
Brabletz T, Pietrowski I, Serfling E 1991. The immunosuppressives FK506
and cyclosporin A inhibit the generation of protein factors binding to the
two purine boxes of the interleukin 2 enhancer. Nucleic Acids Res
19: 61-67.
Bradley BL, Azzawi M, Jacobson M, Assoufi B, Collins JV, Irani AMA,
Schwartz LB, Durham SR, Jeffrey PK, Kay AB 1991. Eosinophils,
T-lymphocytes, mast cells, neutrophils and macrophages in bronchial biopsy
specimens from atopic subjects with asthma: comparison with biopsy
speciments from atopic subjects without asthma and control subjects and
relationship to bronchial hyperresponsiveness. J Allergy Clin
Immunol 88: 661-670.
Broide DH, Gleich GJ, Cuomo AJ, Coburn DA, Federman LB, Schwartz LB,
Wasserman SI 1991. Evidence of ongoing mast cells and eosinophil
degranulation in symptomatic asthma airway. J Allergy Clin Immunol
88: 637-648.
Broide DH, Paine MM, Firestein GS 1992. Eosinophils express interleukin
5 and granulocyte macrophage-colony-stimulating factor mRNA at sites of
allergic inflammation in asthmatics. J Clin Invest 90:
1414-1424.
Brown DA, Nelson FB, Reinherz EL, Diamond DJ 1991. The human
interferon-g gene contains an inducible promoter that can be
transactivated by tax I and II. Eur J Immunol 21: 1879-1885.
Campbell PM, Pimm J, Ramassar V, Halloran PF 1996. Identification of a
calcium-inducible, cyclosporine-sensitive element in the IFN-g promoter
that is a potential NFAT binding site. Transplantation 61:
933-939.
Carter LL, Dutton RW 1996. Type 1 and type 2: a fundamental dichotomoy
for all T-cell subsets. Curr Opin Immunol 8: 336-342.
Casolaro V, Georas SN, Song Z, Zubkoff ID, Abdulkadir SA, Thanos D, Ono
SJ 1995. Inhibition of NF-AT-dependent transcription by NF-kB:
implications for differential gene expression in T helper cell subsets.
Proc Natl Acad Sci USA 92: 11623-11627.
Casolaro V, Georas SN, Song Z, Ono SJ 1996. Biology and genetics of
atopic disease. Curr Opin Immunol 8: 796-803.
Chen L, Oakley MG, Glover JNM, Jain J, Dervan PB, Hogan PG, Rao A,
Verdine GL 1995. Only one of the two DNA-bound orientations of AP-1 found
in solution cooperates with NFATp. Current Biology 5:
882-889.
Choi MSK, Brines RD, Holman MJ, Klaus GGB 1994. Induction of NF-AT in
normal B lymphocytes by anti-immunoglobulin or CD40 ligand in conjunction
with IL-4. Immunity 1: 179-187.
Chytil M, Verdine GL 1996. The Rel family of eukaryotic transcription
factors. Curr Opin Struct Biol 6: 91-100.
Clutterbuck EJ, Sanderson CJ 1988. Human eosinophil hematopoiesis
studied in vitro by means of murine eosinophil differntiation factor
(IL-5): production of functionally active eosinophils from normal human
bone marrow. Blood 71: 646-651.
Cockerill GW, Bert AG, Ryan GR, Gamble JR, Vadas MA, Cockerill PN
1995a. Regulation of granulocyte-macrophage colony-stimulating factor and
E-selectin expression in endothelial cells by cyclosporin A and the T-cell
transcription factor NFAT. Blood 86: 2689-2698.
Cockerill PN, Bert AG, Jenkins F, Ryan GR, Shannon MF, Vadas MA 1995b.
Human granulocyte-macrophage colony-stimulating factor enhancer function
is associated with cooperative interactions between AP-1 and NFATp/c.
Mol Cell Biol 15: 2071-2079.
Coffier E, Joseph D, Vergaftig BB 1991. Activation of guinea pig
eosinophils by human reconbinant IL-5. Selective priming to
platelet-activating factor-acether and interference of its antagonists.
J Immunol 147: 2595-2602.
Crabtree GR 1989. Contingent genetic regulatory events in T lymphocyte
activation. Science 243: 355-361.
Crabtree GR, Clipstone NA 1994. Signal transmission between the plasma
membrane and nucleus of T-lymphocytes. Annu Rev Biochem 63:
1045-1083.
Dent LA, Strath M, Mellor AL, Sanderson CJ 1990. Eosinophilia in
transgenic mice expressing interleukin 5. J Exp Med 172:
1425-1431.
Desai DM, Newton ME, Kadlecek T, Weiss A 1990. Stimulation of the
phosphatidylinositol pathway can induce T-cell activation. Nature
348: 66-69.
Desreumaux P, Bloget F, Seguy D, Capron M, Cortot A, Colombel JF, Janin
A 1996. Interlukin-3, granulocyte macrophage-colony stimulating factor and
interleukin-5 synthesis in eosinophilic gastroenteritis.
Gastroenterology 110: 768-774.
Desreumaux P, Capron M 1996. Eosinophils in alergic reactions. Curr
Opin Immunol 8: 790-795.
Drazen JM, Arm JP, Austen KF 1996. Sorting out the cytokines of asthma.
J Exp Med 183: 1-5.
Dolganov G, Bort S, Lovett M, Burr J, Schubert L, Short D, McGurn M,
Gibson C, Lewis DB 1996. Coexpression of the interleukin-13 and
interleukin-4 genes correlates with their physical linkage in the cytokine
gene cluster on human chromosome 5q23-31. Blood 87:
3316-3326.
Dolmetsch RE, Lewis RS, Goodnow CC, Healy JI 1997. Differential
activation of transcription factors induced by Ca^2+ response amplitude
and duration. Nature 386: 855-858.
Foster PS, Hogan SP, Ramsay AJ, Matthaei KI, Young IG 1995. IL-5
deficiency abolishes eosinophilia, airways hyperreactivity, and lung
damage in a mouse asthma model. J Exp Med 183: 195-201.
Gonzalo JA, Jia GQ, Aguirre V, Friend D, Coyle AJ, Jenkins NA, Lin GS,
Katz H, Lichtman A, Copeland N, Kopf M, Gutierrez-Ramos JC 1996. Mouse
eotaxin expression parallels eosinophil accumulation during lung allergic
inflammation but not restricted to a Th2-type response. Immunity
4: 1-14.
Ho AM, Jain J, Rao A, Hogan PG 1994. Expression of the transcription
factor NFATp in a neuronal cell line and in the murine nervous system.
J Biol Chem 269: 28181-28186.
Ho SN, Thomas DJ, Timmerman LA, Li X, Francke U, Crabtree GR 1995.
NFATc3, a lymphoid-specific NFATc family member that is calcium-regulated
and exhibits distinct DNA binding specificity. J Biol Chem
270: 19898-19907.
Hodge MR, Ranger AM, de la Brousse FC, Hoey T, Grusby M, Glimcher LH
1996. Hyperproliferation and dysregulation of IL-4 expression in
NF-ATp-deficient mice. Immunity 4: 397-405.
Hoey T, Sun Y-L, Williamson K, Xu X 1995. Isolation of two new members
of the NFAT gene family and functional characterization of the NFAT
proteins. Immunity 4: 461-472.
Hutchinson LE, McCloskey MA 1995. FceRI-mediated induction of nuclear
factor of activated T-cells. J Biol Chem 270: 16333-16338.
Jain J, Miner Z, Rao A 1993a. Analysis of the preexisting and nuclear
forms of nuclear factor of activated T cells. J Immunol 151:
837-848.
Jain J, McCaffrey PG, Miner Z, Kerppola TK, Lambert JN, Verdine GL,
Curran T, Rao A 1993b. The T-cell transcription factor NFATp is a
substrate for calcineurin and interacts with Fos and Jun. Nature
365: 352-355.
Jain J, Burgeon E, Badalian TM, Hogan PG, Rao A 1995a. A similar
DNA-binding motif in NFAT family proteins and the Rel homology region.
J Biol Chem 270: 4138-4145.
Jain J, Loh C, Rao A 1995b. Transcriptional regulation of the
interleukin 2 gene. Curr Opin Immunol 7: 333-342.
Jia GQ, Gonzalo JA, Lloyd C, Kremer L, Lu L, Martinez C, Wershil BK,
Gutierrez-Ramos JC 1996. Distinct expression and function of the novel
mouse chemokine monocyte chemotactic protein-5 in lung allergic
inflammation. J Exp Med 184: 1939-1951.
Jose PJ, Griffiths-Johnson DA, Collins PD, Walsh DT, Moqbel R, Totty
NF, Truong O, Hsuan JJ, Williams TJ 1994. Eotaxin: a potent eosinophil
chemoattrac-tant cytokine detected in guinea-pig model of allergic airways
inflammation. J Exp Med 179: 881-887.
Kelly K, Siebenlist U.1995. Immediate-early genes induced by antigen
receptor stimulation. Curr Opin Immunol 7: 327-332.
Kita H and Gleich GJ. 1996. Chemokines active on eosinophils -
potentials roles in allergic inflammation. J Exp Med 183:
2421-2426.
Kopf M, Brombacher F, Hodgkin PD, Ramsay AJ, Milbourne EA, Dai WJ,
Ovington KS, Behm CA, Kohler G, Young IG, Matthaei KI 1996. IL-5-deficient
mice have a development defect in CD5 B-1 cells and lack eosinophilia but
have normal antibody and cytotoxic T cell response. Immunity 4:
15-24.
Leonard DG, Ziff EB, Greene LA 1987. Identification and
characterization of mRNAs regulated by nerve growth factor in PC12
cells. Mol Cell Biol 7: 3156-3167.
Loh C, Shaw KTY, Carew J, Viola JPB, Luo C, Perrino BA, Rao A 1996a.
Calcineurin binds the transcription factor NFAT1 and reversibly regulates
its activity. J Biol Chem 271: 10884-10891.
Loh C, Carew JA, Kim J, Hogan PG, Rao A 1996b. T-cell receptor
stimulation elicits an early phase of activation and a later phase of
deactivation of the transcription factor NFAT1. Mol Cell Biol
16: 3945-3954.
Luo C, Burgeon E, Carew JA, Badalian TM, McCaffrey PG, Lane WS, Hogan
PG, Rao A 1996. Recombinant NFAT1 (NFATp) is regulated by calcineurin in T
cells and mediates the transcription of several cytokine genes. Mol
Cell Biol 16: 3955-3966.
Lopez AF, Begley CG, Williamson DJ, Warren DJ, Vadas MA, Sanderson CJ
1986. Murine eosinophils differentiation factor: an eosinophil-specific
colony-stimulating factor with activity for human cells. J Exp Med
163: 1085-1099.
Mattila PS, Ullman KS, Fiering S, Emmel EA, McCutcheon M, Crabtree GR,
Herzenberg LA 1990. The action of cyclosporin A and FK506 suggest a novel
step in the activation of T lymphocytes. EMBO J 9:
4425-4433.
Masuda E, Naito Y, Tokumitsu H, Campbell D, Saito F, Hannum C, Arai
K-I, Arai N 1995. NFATx, a novel member of the NFAT family that is
expressed predominantly in the thymus. Mol Cell Biol 15:
2697-2706.
McCaffrey PG, Luo C, Kerppola TK, Jain J, Badalian TM, Ho AM, Burgeon
E, Lane WS, Lambert JN, Curran T, Verdine GL, Rao A, Hogan PG 1993.
Isolation of the cyclosporin-sensitive T cell transcription factor
NFATp. Science 262: 750-754.
McCaffrey PG, Goldfeld AE, Rao A 1994. The role of NFATp in cyclosporin
A-sensitive tumor necrosis factor-a gene transcription. J Biol Chem
269: 30445-30450.
Nakajima H, Sano H, Nishimura T, Yoshida S, Iwamoto I 1994. Role of
vascular cell adhesion molecule 1/very late activation antigen 4 and
intercellular adhesion molecule 1/lymphocyte function-associated 1
interactions in antigen-induced eosinophil and T cell recruitment in the
tissue. J Exp Med 179: 1145-1154.
Nolan GP 1994. NF-AT-AP-1, Rel-bZIP: hybrid vigor and binding under the
influence. Cell 77: 795-798.
Northrop JP, Ho SN, Chen L, Thomas DJ, Timmerman LA, Nolan GP, Admon A,
Crabtree GR 1994. NF-AT components define a family of transcription
factors targeted in T-cell activation. Nature 369: 497-502.
Pare PD, Bai TR 1995. The consequences of chronic allergic
inflammation. Thorax 50: 328-332.
Park J, Takeuchi A, Sharma S 1996. Characterization of a new isoform of
the NFAT (nuclear factor of activated T cells) gene family member
NFATc. J Biol Chem 34: 29014-29021.
Paul WE, Seder RA 1994. Lymphocyte responses and cytokines. Cell
76: 241-251.
Perkins D, Wang Z, Donovan C, He H, Mark D, Guan G, Wang Y, Walunas T,
Bluestone J, Listman J, Finn PW 1996. Regulation of CTLA-4 expression
during T cell activation. J Immunol 156: 4154-4159.
Peters SP 1990. Mast cells and histamine in asthma. J Allergy Clin
Immunol 86: 642-646.
Ponath PD, Qin S, Ringler DJ, Clark-Lewis I, Wuang J, Kassam N, Smith
H, Shi X, Gonzalo JA, Newman W et al. 1996. Cloning of the human
eosinophil chemoattractant eotaxin. J Clin Invest 97:
604-612.
Pretolani M, Ruffie C, Silva JRD, Joseph D, Lobb RR, Vergaftig BB 1994.
Antibody to very late activation antigen 4 prevents antigen-induced
bronchial hyperreactivity and cellular infiltration in the guinea pig
airways. J Exp Med 180: 795-805.
Prieschl EE, Gouilleux-Gruart V, Walker C, Harrer NE, Baumruker T
1995a. A nuclear factor of activated T cell-like transcription factor in
mast cells is involved in IL-5 gene regulation after IgE plus antigen
stimulation. J Immunol 154: 6112-6119.
Prieschl EE, Pendl GG, Harrer NE, Baumruker T 1995b. p21ras links FceRI
to NF-AT family member in mast cells: the AP3-like factor in this cell
type is an NF-AT family member. J Immunol 155: 4963-4970.
Randak C, Brabletz T, Hergenröther M, Sobotta I, Serfling E 1990.
Cyclosporin A suppresses the expression of the interleukin 2 gene by
inhibiting the binding of lymphocyte-specific factors to the IL-2
enhancer. EMBO J 9: 2529-2536.
Rao A 1994. NF-ATp: a transcription factor required for the coordinate
induction of several cytokine genes. Immunol. Today 15: 274-281.
Rao A, Luo C, Hogan PG 1997. Transcription factors of the NFAT family:
regulation and function. Ann Rev Immunol 15: 707-747.
Romagnani S 1995. Technological advances and new insights into
pathogenesis prelude novel therapeutic strategies. Curr Opin
Immunol 7: 745-750.
Rooney JW, Sun Y-L, Glimcher LH, Hoey T 1995a. Novel NFAT sites that
mediate activation of the interleukin-2 promoter in response to T-cell
receptor stimulation. Mol Cell Biol 15: 6299-6310.
Rooney JW, Hoey T, Glimcher LH 1995b. Coordinate and cooperative roles
for NFAT and AP-1 in the regulation of the murine IL-4 gene.
Immunity 2: 473-483.
Rothenberg ME, MacLean JA, Pearlman E, Luster AD, Leder P 1997.
Targeted disruption of the chemokine eotaxin partially reduces
antigen-induced tissue eosinophilia. J Exp Med 185: 785-790.
Ruff VA, Leach KL 1995. Direct demonstration of NFATp dephosphorylation
and nuclear localization in activated HT-2 cells using a specific NFATp
polyclonal antibody. J Biol Chem 260: 22602-22607.
Sanderson CJ, Warren DG, Strath M 1985. Identification of a lymphokine
that stimulates eosinophil diffrentiation in vitro. Its relationship to
interleukin 3, and functional properties of eosinophils produced in
cultures. J Exp Med 162: 60-74.
Sarafi MN, Garcia-Zepeta EA, MacLean JA, Charo IF, Luster AD 1997.
Murine monocyte chemoattractant protein (MCP)-5: a novel CC chemokine that
is a structural and functional homologue of human MCP-1. J Exp Med
185: 99-109.
Schubert LA, King G, Cron RQ, Lewis DB, Aruffo A, Hollenbaugh D 1995.
The human gp39 promoter. J Biol Chem 270: 29624-29627.
Sehmi R, Wardlaw AJ, Cromwell O, Kurihara K, Waltmann P, Kay AB 1992.
Interleukin-5 selectively enhances the chemotactic response of eosinophils
obtained from normal but not eosinophilic subjects. Blood
79: 2952-2959.
Serfling E, Avots A, Neumann M 1995. The architecture of the
interleukin-2 promoter: a reflection of T lymphocyte activation.
Biochem Biophys Acta 1263: 181-200.
Shaw JP, Utz PJ, Durand DB, Toole JJ, Emmel EA, Crabtree GR 1988.
Identification of a putative regulator of early T cell activation
genes. Science 241: 202-205.
Shaw KTY, Ho AM, Raghavan A, Kim J, Jain J, Park J, Sharma S, Rao A,
Hogan PG 1995. Immunosuppressive drugs prevent a rapid dephosphorylation
of the transcription factor NFAT1 in stimulated immune cells. Proc Natl
Acad Sci USA 92: 11205-11209.
Sullivan S, Broide DH 1996. Compartimentalization of eosinophil
granulocyte-macrophage colony-stimulating factor expression in patients
with asthma. J Allergy Clin Immunol 97: 966-976.
Takatsu K, Tominaga A, Harada N, Mita S, Matsumoto M, Takahashi T,
Kikuchi Y, Yamaguchi N 1988. T cell-replacing factor (TRF)/interleukin 5
(IL-5): molecular and functional properties. Immunol Rev
102: 107-135.
Tsai EY, Jain J, Pesavento PA, Rao A, Goldfeld AE 1996. Tumor necrosis
factor alpha gene regulation in activated T cells involves ATF-2/Jun and
NFATp. Mol Cell Biol 16: 459-467.
Tsytsykova AV, Tsitsikov EN, Geha RS. 1996 The CD40L promoter contains
nuclear factor of activated T cells-binding motifs which require AP-1
binding for activation of transcription. J Biol Chem 271:
3763-3770.
Ullman KS, Flanagan WM, Edwards CA, Crabtree GR 1991. Activation of
early gene expression in T lymphocytes by Oct-1 and an inducible protein,
OAP40. Science 254: 558-562.
Venkataraman L, Francis DA, Wang Z, Liu J, Rothstein TL, Sen R 1994.
Cyclosporin A-sensitive induction of NF-AT in murine B cells.
Immunity 1: 189-196.
Wagner C, Gundel R, Reily P, Haynes N, Letts L, Rothlein R 1990.
Intercellular adhesion molecule-1 (ICAM-1) in the pathogenesis of asthma.
Science 247: 456-459.
Wang DZ, McCaffrey PG, Rao A 1995. The cyclosporin-sensitive
transcription factor NFATp is expressed in several classes of cells in the
immune system. Ann N Y Acad Sci 766: 182-194.
Wardlaw AJ, Symon FS, Walsh GM 1994. Eosinophil adhesion in allergic
inflammation. J Allergy Clin Immunol 94: 1163-1171.
Warren DJ, Moore MAS 1988. Synergism among interleukin 1, interleukin 3
and interleukin 5 in the production of eosinophils from primitive
hemopoietic stem cells. J Immunol 140: 94-99.
Warringa RAJ, Mengelers HJJ, Kuijper PHM, Raaijmakers JAM, Bruijnzeel
PBL, Koenderman L 1992. In vivo priming of platelet-activating
factor-induced eosinophil chemotaxis in allergic asthmatic individuals.
Blood 79: 1836-1841.
Weiss A, Littman DR 1994. Signal transduction by lymphocyte antigen
receptors. Cell 76: 263-274.
Weiss DL, Hural J, Tara D, Timmerman LA, Henkel G, Brown MA 1996.
Nuclear factor of activated T cells is associated with a mast cell
interleukin 4 transcription complex. Mol Cell Biol 16:
228-235.
Weller PF, Rand TH, Goelz SE, Chi-Rosso G, Lobb RJ 1991. Human
eosinophil adherence to vascular endothelium mediated by binding VCAM-1
and ELAM-1. Proc Natl Acad Sci USA 88: 7430-7435.
Weller PF 1993. Lipid, peptide and cytokine mediators elaborated by
eosinophils, p. 25-42. In H Smith, RM Cook (eds.), Immunopharmacology
of Eosinophils. Academic Press Incorporated, London.
Wu J, Katzav S, Weiss A 1995. A functional T-cell receptor signaling
pathway is required for p95vav activity. Mol Cell Biol 15:
4337-4346.
Yamaguchi Y, Suda T, Suda J, Eguchi M, Miura Y, Harada N, Tominaga A,
Takatsu K 1988. Purified interleukin 5 supports the terminal
differentiation and proliferation of murine eosinophilic precursors. J
Exp Med 167: 43-56.
Yaseen NR, Maizel AL, Wang F, Sharma S 1993. Comparative analysis of
NFAT (nuclear factor of activated T cells) complex in human T and B
lymphocytes. J Biol Chem 268: 14285-14293.
Xanthoudakis S, Viola JPB, Shaw KTY, Luo C, Wallace JD, Bozza PT,
Curran T, Rao A 1996. An enhanced immune response in mice lacking the
transcription factor NFAT1. Science 272: 892-895.
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc97180b.jpg]
[oc97180c.jpg]
[oc97180a.jpg]
|

{kind=link}
{kind=link}
{kind=link}