 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Systemic Modulation of Peripheral Eosinophilia (Air Pouch Model) in Schistosoma mansoni Infection RG Pacheco, HL Lenzi*^/+
Clinica Medica B, Departamento de Medicina Geral, Hospital Universitario
Gaffree e Guinle, Universidade do Rio de Janeiro, Rua Mariz e Barros 775,
20270-020 Rio de Janeiro, RJ, Brasil This work was supported by Fundacao Oswaldo Cruz. Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97182
Sizes of Files:
Text: 32.2K
Graphics: Line drawings and photographs (jpg) - 125K
Tables (jpg) - 27K
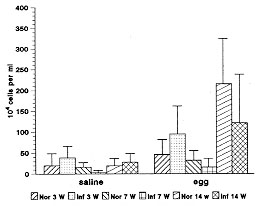
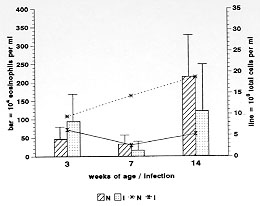
Schistosoma mansoni infection induces in their hosts a marked and sustained eosinophilia, which is influenced or modulated by complex mechanisms, that vary according to the phase of infection. To address this phenomenon, we used the air pouch (AP) model in control and infected Swiss webster mice, analyzing the cellular, tissue response and local expression of adhesion molecules [CD18 (beta2-chain), CD44, ICAM-1 (CD54), L-selectin (CD62L), CD49d (alpha4-chain), LFA1 (CD11a)]. Infected animals were studied at 3 (pre-oviposition phase), 7 (acute phase), and 14 (chronic phase) weeks after infection (5-6 mice/period of infection). Normal mice were age-matched. Results showed that after egg stimulation, compared with matched controls, the infected mice, at each point of infection, showed a lower eosinophil response in the acute (7 weeks) and chronic phase (14 weeks) of infection. However, when the infected mice were in pre-oviposition phase (3 weeks) their eosinophil response surpassed the control ones. In the AP wall of infected mice, a significant decrease in the expression of ICAM-1 and CD44 in fibroblastic-like cells and a reduction in the number of CD18 and CD11a in migratory cells were observed. The other adhesion molecules were negative or weakly expressed. The results indicated that in the air pouch model, in S. mansoni-infected mice: (1) eosinophil response is strikingly down-regulated, during the acute ovular phase; (2) in the pre-oviposition phase, in contrast, it occurs an up-regulatory modulation of eosinophil response, in which the mechanisms are completely unknown; (3) in the chronic phase of the infection, the down modulation of eosinophil response is less pronounced; 4) Down-regulation of adhesion molecules, specially of ICAM-1 appear to be associated with the lower eosinophil response. Key words: air pouch - adhesion molecules - Schistosoma mansoni egg - eosinophil - inflammation Although the eosinophil was described more than one hundred years ago, its actual functions are still not well understood (Wardlaw et al. 1995). A hallmark of some allergic, parasitic, neoplastic, idiopathic and other diseases, eosinophil has been attributed to beneficial as well as harmful functions (Capron 1992, Tagboto 1996). Not withstanding from the ability to kill helminths to the destructive actions in asthma, eosinophil is a complex and mysterious cell. In parasitic diseases, as in schistosomiasis mansoni, eosinophilia is dependent on antigens secreted by eggs through the pores of the egg shell (SEA) (Race et al. 1969, Sher et al. 1990). The bone marrow and the blood eosinophilia peak, respectively, by the fifth and seventh week after the infection, and have been attributed to the enhanced production of interleukin-5. Other primordial precursors stimulating cytokines are also involved in the eosinophilic pathway as the stem cell factor, IL-1, IL-3, GM-CSF, and G-CSF (Clutterbuck et al. 1989, Weller 1992). The eosinophil is one of the most frequent cellular component of the most important lesion of schistosomiasis, the granuloma (Warren 1972). Even present in high number, the eosinophil is not a sine qua non condition for granuloma development (Sher et al. 1990). The acute eosinophil cellular response to eggs, and its contribution to the granuloma formation is not a well studied field. The murine experimental Schistosoma mansoni infection induces, as the infection ages, a modulatory mechanism that lower the hypersensitivity response to eggs (Andrade & Warren 1964, Domingo & Warren 1968, Boros et al. 1975). Schistosomiasis mansoni has been characterized by a chronological change in cytokine expression of Th1 and Th2 subsets (Pearce et al. 1991, Chensue et al. 1993).^ These changes are involved in the modulatory process, and to IL-10 has been attributed a pivotal role in these mechanisms (Stadecker 1994). We have previously shown, in the air pouch model, that S. mansoni egg induces, after 6-8 hr of stimulation, an acute intra air pouch eosinophil wave (Pacheco & Lenzi 1993). In order to get a better insight of the eosinophil participation in schistosomiasis mansoni infection, we performed a sequential study of the eosinophil migration into the air pouch model, after 8 hr of egg stimulation, in mice at 3 (pre-oviposition phase), 7 (acute phase) and 14 (chronic phase) weeks after infection. This study was complemented by an immunofluorescence analysis of the expression of adhesion molecules at the air pouch covering, using monoclonal antibodies. MATERIALS AND METHODS S. mansoni infection - Five days old Swiss webster mice were infected by percutaneous exposure to 70 cercariae of the Belo Horizonte isolate of S. mansoni (Paraense & Correa 1963, 1981). Air pouch - It was prepared as described and reported by Edwards et al. (1981)^ with modifications. Briefly, one day before the beginning of the air pouch production, the fur on the dorsum of each mouse was shaved with an electric clipper. On day zero, 5 ml of sterile air were injected subcutaneously into the same region of mice to form the pouches. In order to keep them patent, on day 3, they were replenished with 2 ml of sterile air. All experimental procedures to the air pouch production were carried out under light ether anesthesia. The sterile air was obtained in a laminar flow station by filtration through a Millipore (0.22 mm) directly into a syringe of 10 ml. On day 8, the pouches were stimulated with eggs or endotoxin-free saline, using individual sterile needles. Egg purification - Mice with 8-12 weeks of infection, as before, were killed by an overdose of ether inhalation. The intestines were collected, opened, water cleaned, chopped and left to stay in a beaker overnight at room temperature. The material was processed in a blender, for a few seconds, in 1.7% saline, filtered through gradually smaller serial size 48, 80, 100, 150 steel mesh sieves and the eggs retained in a size 325 mesh sieve (Bertel, Caieiras, SP, Brasil). The eggs sedimented for one hour and half at 4 C. Finally, the eggs were collected and centrifuged at 150g for 30 seconds, several times until purification. The pellet containing eggs was kept overnight in an ice bath at 0-4 C until use. Injection of eggs - The eggs were adjusted to a 5000/ml concentration in 0.9% endotoxin-free saline and injected by using an insulin needle and syringe. The same number of matched control animals received endotoxin-free saline. Cell harvesting - The animals were killed by an overdose of ether inhalation, and 1 ml of phosphate-buffer saline (PBS) was injected into the air pouches. The pouches were gently massaged and opened, at their caudal portion. The material was collected with a Pasteur pipette and maintained in ice bath until cell counting and cytocentrifugation. Cell counting - Total cell count was done in Neubauer chamber. The absolute eosinophil number was obtained from the percentage of eosinophil in the centrifuged slide, and the total cell count. Cytospin preparation - The collected materials from the air pouches were cytocentrifuged (Cito-spin, Incibras, Brasil) during 10 min, at 640 rpm. The slides were stained by May-Grunwald method, and 200-400 viable cells per slide were counted. Experimental design - Infected Swiss webster mice of both sexes, after 2 (pre-oviposition phase), 6 (acute phase) and 13 (chronic phase) weeks of infection (5-6 mice/group) were picked randomly for the air pouch production. After 8 days, the air pouches were stimulated with 2,500 eggs diluted in endotoxin-free saline; 8 hr later, mice were killed and intrabursal cells were collected. The same number of matched controls which received only endotoxin-free saline were also analyzed. Histopathology - The whole air pouch were fixed in Formalin Millonig (Carson et al. 1973), at pH 7.0-7.4. The samples were embedded in paraffin and the slides stained with hematoxylin/eosin, PAS-Alcian blue pH 1.0 and 2.5, Lennert`s Giemsa, Evans blue for eosinophils detection using confocal microscopy (Lenzi et al. 1996) , in LSM-410 (Zeiss). Direct immunofluorescence - Adhesion molecules - A small piece of the cephalic portion of the air pouch attached to the skin, was immediately frozen in liquid nitrogen and maintained at -20 C until the cryostat slides preparation. The slides were fixed with cold acetone and conserved at -20 C until the immunofluorescence studies. Monoclonal antibodies (PharMingen) diluted in a 1:15 ratio to ICAM-1 (CD54), CD44, CD18 (beta2 chain), LFA1 (CD11a), L-Selectin (CD62L), LEPAM (CD49d) were used. Briefly after rehydration with PBS, the slides were blocked with a solution of 1% BSA and 3 % powder skimmed milk. The diluted mAb was maintained in contact with slides during 45 min at 37 C. The slides were stained, mounted in glycerol with p-phenylenediamine and cover glass and examined in the same day in a microscope for immunofluorescence (Zeiss-Germany). Data analysis - The data are summarized as mean +/- SD, and were analyzed by the statistical GraphPad InStat software package, version 2.0 (Graph Pad Software, San Diego, CA, USA). Statistical significance was determined by using unpaired t-test and significance was determined with the use of p values < 0.05. RESULTS Absolute eosinophil number - Saline stimulation elicited the lowest eosinophil responses, even in infected mice. After egg stimulation, compared with matched controls, the infected mice, in each point of infection, showed a lower eosinophil response in the acute (7 weeks) and chronic phase (14 weeks) of infection. However, when the infected mice were in pre-oviposition phase (3 weeks), their eosinophil response surpassed the control ones (Fig. 1). Although the eosinophil levels decreased in normal and infected mice, at the 7th week of age/infection, the normal mice showed a continuous increase of total cell count during the experiment. Otherwise, the infected ones presented always lower total cell count than the controls, specially at the 7th week of infection ( Fig. 2)
Figure 2: total cell count (line) and absolute (bar) counts in the air pouches of normal (N) and Schistosoma mansoni-infected-mice (I), 8 hr after S. mansoni egg stimulation, during different times of age/infection.
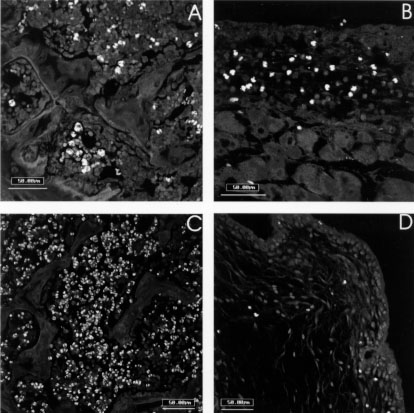
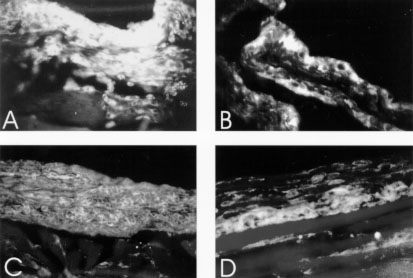
L-selectin (CD62L), CD49d (integrin alpha4-chain) and CD11a (LFA-1) did not show significative difference between infected and control mice, and no one correlation could be drawn relating to the eosinophil migration. CD18 labeled weakly fibroblasts of the air pouch wall of all groups, and was more intense on migrating cells, which were more numerous in air pouch of control mice stimulated with eggs (Fig. 5A). CD54 (ICAM-1), after egg stimulation, marked diffuse and intensely all the air pouch wall layers of the control mice (Fig. 5B), showing impressive decrease in infected ones. This different behavior of CD54 expression was also detected in vascular endothelium of arteries, veins and capillaries. CD44 showed similar staining characteristics as CD54 (ICAM-1), except that it was more intense on fibroblasts and in the innermost layers of the air pouch wall (Fig. 5C, D). Frequently, CD54 and CD44 presented a cadherin-like pattern of immunofluorescence (Table). The lowest expression of CD18, CD54, and CD44 was coincident with the decrease in the number of eosinophils inside the air pouch.
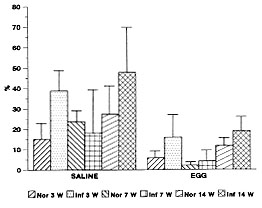
This study showed that S. mansoni infected mice modulated the acute eosinophil response, depending on the time of the infection. This phenomenon was observed using the air pouch model stimulated with purified S. mansoni eggs. In the pre-oviposition phase of the infection occurred an up-modulation, while in the acute (7 weeks) and chronic (14 weeks) phases, the eosinophil response was down-modulated. The literature has emphasized multiple putative immunoregulatory mechanisms, during the course of S. mansoni infection, such as suppressor T cell activity (Colley et al. 1978), adherent phagocytic suppressor cells (Todd et al. 1979), serum-mediated suppression (Colley et al. 1977, Ottesen & Poindexter 1980), circulating immune complexes (Goes et al. 1991) and idiotypic/anti-idiotypic interactions, involving also anti-idiotypic T lymphocytes (Lima et al. 1986, Powell & Colley 1987, Parra et al. 1988, 1991). However, all these mechanisms are related to the pos-oviposition phases of the infection, and probably some of them interfere with eosinophil response. Authors (Colley 1972, Mahmoud et al. 1975, Lenzi & Lenzi 1990) have shown that the eosinophil number fluctuate, during the S. mansoni infection, in distinct compartments (bone marrow, blood, peritoneal cavity, milky spots and organs with egg embolism), reaching higher number in the acute phase, with high levels persisting even in chronic phase of infection. Our results showed that the down modulation of the intrabursal eosinophil response was coincident with the largest systemic and peri-ovular eosinophilia (around 7 weeks), indicating, two important aspects: (1) The down-modulation is independent of eosinophil production and circulation, not being due to a lack of eosinophils supply; (2) It is a striking modulation because it occurs when the peripheral and central (bone marrow) eosinophil levels are very high (Fig. 4C ). The data also suggest that the modulatory mechanisms act, at least, on two levels: endothelium, and air pouch wall. This hypothesis was based on the decrease of the following adhesion molecules: (a) ICAM-1 in vascular endothelium of arteries, veins and capillaries that are adjacent to the air pouch wall and (b) ICAM-1 and CD44 in the air pouch wall. These events could interfere with leukocyte-endothelial recognition phase of the acute inflammatory response, decreasing the number of migrating cells, detected by anti-CD18. Other additional mechanisms can contribute to explain the down-modulation in the acute phase: (1) massive and compartmentalized mobilization of blood eosinophils to organs that suffer egg embolism; (2) large release of soluble adhesion molecules to the circulation, blocking leukocyte-endothelial interactions (Evan Secor et al. 1994); (3) compartmentalized response of the air pouch to egg stimulation. Using protein A-stimulated air pouch, Teixeira et al. (1994) showed a decrease of polimorphonuclear migration in mice infected with S. mansoni, mainly in the acute phase of the infection. The same phenomenon was observed by Abath et al. (1988), in acute Trypanosoma cruzi infected mice; and (4) alterations of eosinophil capabilities by soluble factors. Dessein et al. (1984) showed that the "Eosinophil Cytotoxicity Enhancing Activity" (ECEA) released by blood mononuclear cells was suppressed in most patients, except in those with heavy Schistosoma infections, including patients with hepatosplenomegaly. As is shown in Fig. 1, when the intrabursal eosinophilia of control versus infected mice was compared in post-oviposition phases, always the infected groups attained lower number of eosinophils by cytological analysis. However, when the eosinophil levels from animals with 7 weeks were compared with ones of 14 weeks of infection, there was an absolute and relative increase in the later ones (Figs 1 and 3). This event suggest the existence of different and unknown control mechanisms on the inflammatory reaction in the acute and chronic phases of the schistosomal infection. Lenzi (1991) has shown that in the chronic phase of infection, when compared with the acute one, occurred a decrease of eosinopoiesis, mega-karyopoiesis, and lymphopoiesis, and an increase in the tissue mast and plasma cells, indicating changes in growth factors and cytokines networks during the course of the infection. Our data also showed an enhance of the eosinophil migratory capability to the air pouch, with advancing age, better observed in control group, by the increase of the eosinophil percentage, after saline stimulation (Fig. 3). The age and chronic-infected-related changes do not involve defects in the functions of T cells, including cytokine production (Engwerda et al. 1996). Actually, the studied animals were in the range of young adults (Hobbs et al. 1993). It is interesting to note that, in the infected mice, the eosinophil response inside the air pouch was inversely related to the systemic eosinophil supply. The up-modulation of the intrabursal eosinophilia detected in the pre-oviposition phase of S. mansoni infection, could be due to lack of immune modulation that is part of the acute syndrome (Cheever 1992). Indeed, the first peak of blood eosinophilia that occurs in the pre-oviposition phase is not affected by T-lymphocyte depletion (Colley et al. 1973, Fine et al. 1973, Lenzi et al 1987). . In conclusion, the results of this study, using the air pouch model, showed that the acute eosinophil response is differently modulated, according to the time of S. mansoni infection. Bursal (air pouch) and endothelial cell-derived adhesion molecules, such as ICAM-1 (and CD44 ?) appear to play a prominent role. The egg is the main factor that switch, not only the immune, but also the acute inflammatory response. Further comprehensive studies using intercrines a (C-X-C), and b (C-C) subfamilies (Oppenheim et al. 1991), affiliated with the recruitment phase of the inflammatory response should be applied to a better elucidation of the acute tissue eosinophilia. REFERENCES
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97182f.jpg] [oc97182b.jpg] [oc97182d.jpg] [oc97182a.jpg] [oc97182e.jpg] [oc97182c.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}