 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
A Role for Lymphocytes and Cytokines on the Eosinophil Migration Induced by LPS
Hugo C Castro-Faria-Neto^+, Carmen M Penido, Andrea P Larangeira,
Nucleo de Imunofarmacologia, Departamento de Fisiologia e Farmacodinamica,
Instituto Oswaldo Cruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ,
Brasil. Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97186
Sizes of Files:
Text: 21K
Graphics: Line drawings and photographs (jpg) - 16.8K
In the present work we review the existing evidence for a LPS-induced cytokine-mediated eosinophil accumulation in a model of acute inflammation. Intrathoracic administration of LPS into rodents (mice, rats or guinea pigs) induces a significant increase in the number of eosinophils recovered from the pleural fluid 24 hr later. This phenomenon is preceded by a neutrophil influx and accompanied by lymphocyte and monocyte accumulation. The eosinophil accumulation induced by LPS is not affected by inhibitors of cyclo or lipoxygenase nor by PAF antagonists but can be blocked by dexamethasone or the protein synthesis inhibitor cycloheximide. Transfer of cell-free pleural wash from LPS injected rats (LPS-PW) to naive recipient animals induces a selective eosinophil accumulation within 24 hr. The eosinophilotactic activity present on the LPS-PW has a molecular weight ranging between 10 and 50 kDa and its effect is abolished by trypsin digestion of the pleural wash indicating the proteic nature of this activity. The production of the eosinophilotactic activity depends on the interaction between macrophages and T-lymphocytes and its effect can not be blocked by anti-IL-5 monoclonal antibodies. Accumulated evidence suggest that the eosinophil accumulation induced by LPS is a consequence of a eosinophilotactic cytokine produced through macrophage and T-cell interactions in the site of a LPS-induced inflammatory reaction. Key words: LPS - eosinophil - macrophages - T-cells - cytokine Pleural inflammation induced by LPS LPS is classically known as capable of inducing an inflammatory reaction when injected locally in the tissues. This phenomenon is characterized by an early neutrophil influx followed by a late mononuclear cell accumulation (Issekutz et al. 1987). We have previously demonstrated that the injection of LPS in the pleural cavity of rats, mice or guinea pigs induces an acute (4 hr) neutrophil accumulation that can be inhibited by pretreatment with PAF antagonists. Simultaneously, a decrease in the number of neutrophils in the bone marrow and an increase in blood neutrophils was observed, indicating that neutrophils were being mobilized from the bone marrow pool of cells by LPS stimulation (Bozza et al. 1993b). In addition, eosinophils and mononuclear cells also accumulated in the pleural cavity of LPS injected animals after 24 hr. The number of those cells remained above control values for at least 96 hr (Bozza et al. 1991). Despite many attempts using different doses of LPS we could not detect increased vascular permeability, i.e. plasma leakage, in any of the time points analyzed. It is interesting to point out that until that work, the ability of LPS in inducing eosinophil accumulation had not been studied thoroughly. Actually, only Folkerts et al. (1988) had detected an increase in eosinophil numbers in the brochoalveolar lavage of LPS stimulated guinea pigs. This fact led us to further investigate the mechanism by which LPS induces eosinophil accumulation. Pharmacological modulation of LPS-induced eosinophil accumulation The eosinophil accumulation induced by LPS is not a consequence of a direct chemotactic effect of LPS upon eosinophils since LPS is not able to induce eosinophil migration in vitro (Bozza et al. 1991). This led us to investigate the involvement of endogenous mediators that could account for the eosinophil accumulation observed after LPS challenge. Several mediators can trigger eosinophil accumulation in the course of an acute inflammatory response. Among those, PAF, LTB4 (Silva et al. 1991) and bradykinin (Pasquale et al. 1991) were shown to induce delayed and long-lasting eosinophil infiltration in the rat pleural cavity. Nevertheless, none of these mediators seem to be involved on LPS-induced eosinophil accumulation (Bozza et al. 1993a). Pretreatment with a cyclooxygenase inhibitor (indomethacin), a dual cyclo/lipoxygenase (BW755C), a more specific lipoxygenase inhibitor (BWA4C), PAF receptor antagonists (PCA 4248 and WEB 2086), a combination of BWA4C and WEB 2086, or a bradykinin B2 receptor antagonist (HOE 140) failed to diminish the eosinophil influx induced by LPS (Bozza et al. 1993a; unpublished results). By contrast, dexamethasone and the protein synthesis inhibitor, cycloheximide, were able to abolish LPS-induced eosinophil accumulation (Bozza et al. 1993a). Dexamethasone has different effects that could explain its inhibition of LPS-induced eosinophil accumulation. For instance, dexamethasone is able to induce the synthesis of lipocortins with consequent inhibition of phospholipase A2 and thus inhibition of eicosanoids and PAF synthesis. Nevertheless, in view of the above mentioned results using selective blockers of eicosanoid synthesis and PAF receptor, it is unlikely that this action of dexamethasone is accounting for the inhibition of eosinophil accumulation. Dexamethasone can also directly inhibit the transcription of some genes coding for cytokines such as GM-CSF and IL-3, both able to induce eosinophil activation (Owen et al. 1987, Rothenberg et al. 1989). Together with the results obtained with the protein synthesis inhibitor cycloheximide, which also inhibited LPS-induced eosinophil accumulation, we can further suggest that the increase in the number of pleural eosinophils following LPS stimulation requires the synthesis of a proteic factor, probably a cytokine. Detection of eosinophilotactic activity in the pleural wash of LPS-injected animals Based on the results obtained with the pharmacological experiments, we aimed to clarify whether a protein with eosinophilotactic activity has been produced and released in the inflammatory fluid of LPS-injected animals. Firstly, we demonstrated that the cell-free pleural wash of LPS-injected animals (LPS-PW) is able to induce eosinophil accumulation when injected intrathoracically in recipient naive animals or in animals pretreated with dexamethasone or cycloheximide (Bozza et al. 1993a). These results indicate that a factor with eosinophil chemoattractant activity is released in the pleural cavity after LPS stimulation. Furthermore, these results also indicate that the eosinophilotactic activity in the pleural wash is not the result of LPS contamination since dexamethasone and cycloheximide inhibit eosinophil accumulation induced by LPS itself but do not affect the one induced by the injection of LPS-PW. In contrast to other known eosinophil acting cytokines such as IL-3 (Ulich et al. 1989), TNF (Ulich et al. 1991), IL-8 (Collins et al. 1993) and IL-5 (Warren & Moore 1988), the eosinophilotactic activity in the LPS-PW could not be destroyed by 30 min boiling indicating that this activity is heat-stable. Nevertheless, completely loss of activity was obtained when LPS-PW was incubated with trypsin for 30 min at 37 C, adding support to the interpretation that the activity resides on a newly synthesized protein. Ultrafiltration of LPS-PW showed that this protein has a molecular weight in the range of 10 to 50 kDa (Bozza et al. 1993a). It is important to note that, although IL-5 is able to induce eosinophil accumulation in the pleural cavity and seems to account for the eosinophilia observed in allergic inflammatory reactions (Kaneko et al. 1991, Bozza et al. 1994), incubation of LPS-PW with anti-IL-5 monoclonal antibodies did not inhibited the eosinophil attractant activity present in the pleural wash showing that an eosinophilotactic factor different from IL-5 is responsible for the activity in the LPS-PW. Furthermore, semi-purified LPS-PW was unable to displace the binding of ^125I - RANTES to isolated guinea pigs eosinophils (Bozza et al. unpublished observations) indicating that the activity in LPS-PW is different from the eosinophilotactic chemokine RANTES as well. Other eosinophil attractant chemokines such as MCP-2 (Chang et al. 1989), MCP-3 (Van Damme et al. 1992), and eotaxin (Jose et al. 1994) have been identified but their involvement on LPS-induced eosinophil accumulation remains to be determined. Cellular source of the eosinophilotactic activity present in the LPS-PW Cells accumulating in the site of an inflammatory reaction can contribute to its development by secreting inflammatory mediators such as bioactive lipids, peptides, amines and proteins. Therefore, we hypothesized that one of the resident cells or cells infiltrating the pleural cavity during the initial stages of LPS-induced inflammation could be involved in the production of the factor responsible for the late eosinophil accumulation observed. In this respect, we have shown (Bozza et al. 1994) that the depletion of neutrophils or platelets by the use of specific antibodies did not affect the eosinophil influx induced by LPS. Also, the induction of mast cell degranulation by pre-exposure to low doses of polymixin B could not interfere with the phenomenon. It is important to note, that all those three cell populations are potential sources of eosinophil chemotactic factors since they are able to produce and/or secrete IL-8, RANTES and ECF-A, respectively (Schwartz 1987, Striter et al. 1990, Kameyoshi et al. 1992).
T-lymphocytes are also another major source of eosinophil chemoattractant
cytokines. Those cells are generally believed to mediate the eosinophil
accumulation observed during allergic reactions and parasitic infections
through a mechanism dependent on IL-5 (Spry 1988, Frew et al. 1989, Iwamoto
et al. 1992). To investigate the putative involvement of lymphocytes on the
eosinophil accumulation triggered by LPS the animals were pretreated with
cyclosporin A, an inhibitor of T-lymphocyte activation (Borel 1989). Under
this condition, LPS-induced eosinophil accumulation was drastically reduced
suggesting that lymphocytes might play at least a partial role in the
phenomenon. This result was confirmed by the depletion of T-lymphocytes
using an anti-Thy 1.0 monoclonal antibody which also decreased LPS-induced
eosinophil accumulation (Bozza et al. 1994). We have observed that
T-lymphocytes of the gd^+-subtype infiltrate in the pleural cavity after
LPS stimulation whereas CD4^+ T-cell and CD8^+ T-cell subsets did not.
Depletion of gd^+ T-cell subset with a monoclonal antibody, but not the
depletion of CD4^+ T-cell or CD8^+ T-cell subsets, markedly reduced the
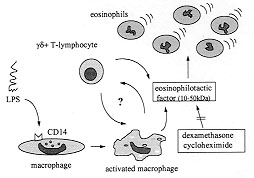
eosinophil accumulation suggesting that gd^+ Macrophages and monocytes express high amounts of the LPS receptor, CD14, and are able to produce and release a variety of cytokines and chemokines upon LPS stimulation (Rietchel & Blade 1992, Ziegler-Heitbrock & Ulevitch 1993, Meheus et al. 1993). These features make resident macrophages good candidates as source of the eosinophilotactic activity detected in the LPS-PW. To investigate this possibility resident pleural macrophages were depleted by means of an intrapleural injection of liposomes containing dichloromethylene diphosphonate. We have previously demonstrated that this procedure was able to induce a selective depletion of pleural macrophages within five days (Bozza et al. 1994) and has no effect on other cells at the same time. In fact, when the animals were pretreated with liposome-entrapped dichloromethylene diphosphonate an inhibition of LPS-induced eosinophil accumulation was observed. This inhibition was not observed when LPS-PW, instead of LPS, was injected into macrophage-depleted animals, indicating that resident macrophages are involved in the production of the eosinophilotactic activity found on LPS-PW rather then contributing to its effect. Moreover, the injection of the supernatant recovered from monolayers of cultured pleural macrophages stimulated by LPS was able to induce eosinophil accumulation in naive animals (Bozza et al. 1994) which reinforce the hypothesis mentioned above. In summary, the eosinophil accumulation induced by LPS appears to involve the synthesis of a protein with molecular weight between 10 and 50 kDa. The presence of T-lymphocytes and macrophages seems to be required to the full development of the eosinophil response to LPS (Fig.). Whether this protein is one of the known eosinophilotactic factors already described or a new cytokine with eosinophil chemoattractant activity will await the isolation and sequence determination of this eosinophilotactic protein.
To Dr MGMO Henriques, Dr MA Martins, Dr PMR Silva, Dr RR Santos and Dr RSB Cordeiro whose contributions were important in different parts of this work. PTB is a PEW fellow in biomedical research. This work was supported by PAPES, CAPES, CNPq and FAPERJ. References
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc97186a.jpg] |
| |||||||||

{kind=link}