
|
Memórias do Instituto Oswaldo Cruz
Fundação Oswaldo Cruz, Fiocruz
ISSN: 1678-8060 EISSN: 1678-8060
Vol. 92, Num. s2, 1997, pp. 205-210
|
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 92 (Suppl.II), 1997, pp.
205-210
Participation of Interleukin-5, Interleukin-8 and Leukotriene B4 in
Eosinophil Accumulation in Two Different Experimental Models
Sandra HP Oliveira, Lucia H Faccioli, Sergio H Ferreira, Fernando Q Cunha^+
Departamento de Farmacologia, Faculdade de Medicina de Ribeirao Preto,
Universidade de Sao Paulo, Av. Bandeirantes 3900, 14049-900 Ribeirao Preto,
SP, Brasil
^+Corresponding author. Fax.: + 55-16-633.2301. E-mail: fdqcunha@fmrp.usp.br
Received 3 September 1997; Accepted 30 September 1997
Code Number:OC97188
Sizes of Files:
Text: 27.2K
Graphics: Line drawings and photographs (jpg) - 105.1K
There are several experimental models describing in vivo
eosinophil (EO) migration, including ip injection of a large volume of
saline (SAL) or Sephadex beads (SEP). The aim of this study was to
investigate the mechanisms involved in the EO migration in these two
models. Two consecutive injections of SAL given 48 hr apart, induced a
selective recruitment of EO into peritoneal cavity of rats, which
peaked 48 hr after the last injection. SEP, when injected ip, promoted
EO accumulation in rats. The phenomenom was dose-related and peaked 48 hr
after SEP injection. To investigate the mediators involved in this process
we showed that BW A4C, MK 886 and dexamethasone (DXA) inhibited the EO
migration induced by SAL and SEP. To investigate the source of the EO
chemotactic factor we showed that mast cells, macrophages (MO), but not
lymphocytes, incubated in vitro in presence of SAL released a factor
which induced EO migration. With SEP, only mast cells release a factor that
induced EO migration, which was inhibited by BW A4C, MK 886 and DXA.
Furthermore, the chemotactic activity of SAL-stimulated mast cells was
inhibited by antisera against IL-5 and IL-8 (interleukin). SAL-stimulated
MO were only inhibited by anti-IL-8 antibodies as well SEP-stimulated mast
cells. These results suggest that the EO migration induced by SAL may be
dependent on resident mast cells and MO and mediated by LTB4, IL-5 and
IL-8. SEP-induced EO migration was dependent on mast cells and may be
mediated by LTB4 and IL-8. Furthermore, IL-5 and IL-8 induced EO migration,
which was also dependent on resident cells and mediated by LTB4 . In
conclusion, EO migration induced by SAL is dependent on mast cells and MO,
whereas that induced by SEP is dependent on mast cells alone. Stimulated
mast cells release LTB4, IL-5 and IL-8 while MO release LTB4 and IL-8. The
IL-5 and IL-8 release by the SAL or SEP-stimulated resident cells may act
in an autocrine fashion, thus potentiating LTB4 release.
Key words: interleukin-5 - interleukin-8 - leukotrine B4 -
eosinophil migration - saline - Sephadex
Participation of Interleukin-5, Interleukin-8 and Leukotriene B4 in
Eosinophil Accumulation in Two Different Experimental Models (Full
Text)
Eosinophils are thought to play an important role in many inflammatory and
allergic diseases such as asthma (Barnes et al. 1988, Gleich 1990), atopic
dermatitis (Leiferman et al. 1985), allergic rhinitis (Bascon et al. 1989)
and parasitic infections (Kay 1985). Although eosinophils are involved in
host defense mechanisms against parasites (Capron 1992), they can cause
damage to mammalian tissues through a variety of mechanisms, including the
release of granule-derived cytotoxic proteins (Gleich et al. 1988) and the
generation of toxic oxygen radicals (Davies et al. 1984). Thus,
understanding the mediators involved in eosinophil migration may allow us
to develop procedures for treating various pathological processes. Several
experimental models describing eosinophil migration into the extravascular
space have been reported. These models include the injection of polymyxin B
(Sun et al. 1985), alum adjuvant (Walls 1977), antigen-coated latex
(Schriber & Zucker-Franklin 1974), parasitic larvae or their extracts
(Auriault et al. 1983) and large volumes of physiological saline (Cook et
al. 1987). However, the endogenous mediators responsible for eosinophil
recruitment to the site of inflammation have not yet been fully
characterized. Furthermore, intravenous injection of Sephadex has been
shown to induce blood and lung eosinophilia in rats and may contribute to
bronchial hyperreactivity in vivo (in rats) and ex vivo (in
guinea pigs) (Spicer et al. 1990, Maghni et al. 1993). The mechanisms by
which these beads induce eosinophilia also need to be further elucidated.
In addition, several factors including leuko-triene B4 (LTB4) (Hakansson et
al. 1987, Faccioli et al. 1991), C5a (Ogawa et al. 1981), PAF (Czarnetzki &
Csato 1989), interleukin-5
(IL-5) (Lopez et al. 1988, Sehmi et al.
1992), interleukin-2 (IL-2) (Rand et al. 1991, Meacock et al. 1991),
interleukin-8 (IL-8) (Collins et al. 1993), Rantes (Kameyoshi et al. 1992),
eotaxin (Jose et al. 1994) and factors derived from mast cells (Holgate
1991, Raible et al. 1992), lymphocytes (Berman & Weller 1992) and
macrophages (Lee & Lane 1992) are known to induce eosinophil migration
in vivo and in vitro. LTB4, IL-5, IL-8 and PAF have been also
detected at the site of inflammatory and allergic reactions (Resnick &
Weller 1993). However, it is not yet established whether these mediators
are direct chemoattractants or act indirectly by stimulating the release of
other eosinophil chemotactic factors from resident cells.
The aim of this study was to investigate the mediators involved in
eosinophil migration induced by a large volume of saline and by Sephadex
beads and the mechanisms by which these mediators induce eosinophil
recruitment to the peritoneal cavity of rats.
Figure 1: induction
of eosinophil migration by
injection of large volumes of saline or Sephadex into the peritoneal cavity
of rats. Time-course of the eosinophil migration induced by two consecutive
injections of 5 ml PBS (o)>; control), saline (SAL, panel a, triangle) or a
single injection of 2 ml Sephadex (SEP, panel c, solid circle) and
dose-response curve of eosinophil migration induced by saline (panel b) or
Sephadex (panel d) injection into rat peritoneal cavities evaluated 48 hr
after the injection of Sephadex or after the last injection of saline. The
results are presented as means +/- SEM for six animals per group. The
asterisk indicates a significant difference between the saline or Sephadex
and PBS groups (p<0.05; ANOVA followed by Bonfferoni's t test).
Figure 1a shows that two
consecutive injections of saline
(SAL: 0.15 M) at 48 hr intervals induced significant eosinophil migration
into the peritoneal cavity of naïve rats 48 hr after the second injection.
Eosinophil migration was not influenced by increasing concentrations of
sodium chloride (Fig. 1b). Thus, the concentration of SAL used in
subsequent experiments was fixed at 0.15 M. In contrast to saline, PBS did
not promote eosinophil migration. Therefore, to investigate the mechanism
by which saline induced eosinophil migration, we added various ions to SAL
in order to reconstitute the PBS ion concentration. The addition of
potassium chloride (but not phosphate) blocked the ability of SAL to induce
eosinophil migration as shown by the eosinophils/ml of peritoneal wash
fluid (means +/- SEM): control (PBS), 0.17 +/- 0.05x10^6; SAL, 1.1 +/-
0.06x10^6; SAL+ phosphate, 1.4 +/- 0.3x10^6; SAL + potassium chloride, 0.3
+/- 0.1x10^6* and SAL+ potassium chloride + phosphate, 0.4 +/- 0.1x10^6 (*p
< 0.05, n=6, Student's t-test). A similar inhibition of the ability of SAL
to induce the in vitro release of a chemotactic factor for
eosinophils by mast cells or macrophages (see bellow) was seen by
correcting the potassium concentration to the corresponding values for PBS.
The mechanism by which SAL induced the in vitro release of the
eosinophil chemotactic factors or in vivo eosinophil migration may
involve an alteration of cell membrane properties resulting from a decrease
in the potassium ion concentration in the extracellular environment.
Potassium chloride is known to play a role in the control of the resting
membrane potential (Cook 1988, Janiszewski et al. 1992).
With Sephadex, it was observed that intraperitoneal injection of this
stimulus promoted time-dependent eosinophil accumulation (Fig. 1c, d).
To investigate the mediators involved in eosinophil migration induced by
saline and Sephadex, the effect of anti-inflammatory drugs on the
eosinophil migration induced by both stimuli was evaluated. Pretreatment of
the animals with a PAF antagonist (BN 52021, 20 mg/kg/day) or a
cyclooxygenase inhibitor (indomethacin, 5 mg/kg/day) had no effect on
eosinophil migration while the 5-lipoxygenase inhibitors BW A4C (20
mg/kg/day) and MK 886 (1 mg/kg/day) and the glucocorticoid, dexamethasone
(0.5 mg/kg/day) inhibited migration induced by either stimulus ( Fig. 2). Thus, PAF-acether and
cyclooxygenase products seem not
to be involved in SAL or Sephadex-induced eosinophil migration since BN
52021 or indomethacin did not affect the response. The mediator involved in
eosinophil migration induced by SAL or Sephadex appears to be a
5-lipoxygenase product since the migration was inhibited by pretreatment of
the animals with BW A4C and MK 886. LTB4 is a plausible candidate since it
induced eosinophil migration in our model (data not shown), as well as in
an in vitro assay. In this context, the association between
hypersensitivity reactions, the presence of eosinophils and the detection
of LTB4 in lung tissue has been recently suggested.
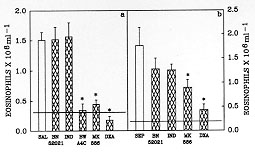
Figure 2: BW A4C, MK
886 and dexamethasone, but not BN
52021 or indomethacin, inhibited the in vivo eosinophil migration
induced by saline or Sephadex. The open bar shows the eosinophil migration
induced by two injections of saline (SAL, panel a) or Sephadex (SEP, panel
b) into PBS-pretreated animals. The hatched bars represent the eosinophil
migration in rats pretreated with BN 52021 (20 mg/kg/day), indomethacin
(IND, 5 mg/kg/day), BW A4C (20 mg/kg/day), MK 886 (MK, 1 mg/kg/day) and
dexamethasone (DXA, 0.5 mg/kg/day). Eosinophil migration was evaluated 48
hr after the injection of SEP or after the last injection of SAL. The
dashed line represents the number of eosinophils in rats injected twice
with PBS. The results are presented as means +/- SEM for six animals per
group. The asterisk indicates significant inhibition compared to the
response with saline or Sephadex (p<0.05; ANOVA followed by Bonfferoni's t
test).
In order to investigate the role of resident peritoneal cells in SAL- or
Sephadex-induced eosinophil migration, isolated peritoneal mast cells,
peritoneal macrophages, or lymphocytes collected from the thoracic duct
were preincubated with PBS, SAL or Sephadex and the ability of the
supernatants to induce eosinophil migration was tested. The supernatants of
mast cells and macrophages incubated with SAL, but not with PBS, induced
significant eosinophil migration 6, 24 and 48 hr after injection into the
peritoneal cavities of naïve rats. In contrast, the supernatant of
lymphocytes incubated with SAL was unable to induce eosinophil migration
(Fig. 3a). In the Sephadex
model, only the supernatant from
Sephadex-stimulated mast cells induced significant eosinophil migration
when injected into the peritoneal cavities of naïve rats (Fig. 3b). These
data suggest that the eosinophil migration induced by SAL is dependent on
resident macrophages and/or mast cells whereas that induced by Sephadex
only depends on mast cells.
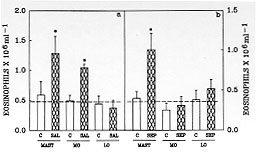
Figure 3: rat
peritoneal resident cells release an
eosinophil chemotactic factor when incubated with saline or Sephadex. Mast
cells and macrophages, but not lymphocytes, incubated with saline (panel a)
released an eosinophil chemotactic factor into the supernatant. Mast cells
but not macrophages or lymphocytes incubated with Sephadex (panel b)
released an eosinophil chemotactic factor into the supernatant. The
supernatants were ultrafiltered and resuspended in the same volume of PBS.
Eosinophil migration was evaluated 6 hr after injection of the supernatant.
The dashed line represents the number of eosinophils in rats injected with
PBS. The results are presented as means +/- SEM for six animals per group.
The asterisk indicates significant differences in eosinophil migration
induced by SAL or SEP and PBS supernatants (p<0.05; ANOVA followed by
Bonfferoni's t test).
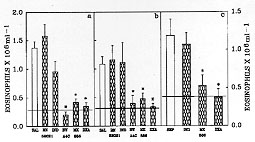
Figure 4: BW A4C, MK
886 and dexamethasone, but not BN
52021 or indomethacin, inhibited the release of eosinophil chemotactic
factor by mast cells and macrophages incubated with saline or by mast cells
incubated with sephadex. The bars represent the eosinophil migration
induced by the injection of 3 ml of the supernatant of mast cells (a) or
macrophages (b) stimulated with saline alone (open bars) or supernatant of
mast cells (c) stimulated with Sephadex alone (open bars). The hatched bars
represent the cells stimulated with saline or Sephadex plus BN 52021 (100
uM), indomethacin (10 mM), BW A4C (100 uM), MK 886 (1 uM) or dexamethasone
(10 uM). The dashed line represents the number of eosinophils after the
injection of 3 ml of PBS alone (control). The results are presented as
means +/- SEM for six animals per group. The asterisk indicates significant
differences between the group incubated with saline or Sephadex alone and
the groups treated with various drugs (p<0.05; ANOVA followed by
Bonfferoni's t test).
The pretreatment of mast cells or macrophages with BW A4C (100 uM), MK 886
(1 uM) or dexamethasone (10 uM), but not with indomethacin (10 uM) or BN
52021 (100 uM), inhibited the release of the eosinophil chemotactic factor
into the supernatants of these cells stimulated by SAL (Fig. 4a,
b). Together with the in vivo experiments, these results suggest
that eosinophil migration induced by saline is mediated by LTB4, which is
released by the resident mast cells and macrophages. In the Sephadex model,
the pretreatment of mast cells with MK 886 (1 uM) or dexamethasone (10 uM),
but not with indomethacin (10 uM), inhibited the release of the eosinophil
chemotactic factor into the supernatant of mast cells stimulated by
Sephadex (Fig. 4c). Together with the in vivo data, these results
support the suggestion that LTB4 is an important mediator of SAL- or
Sephadex-induced eosinophil migration. To investigate whether cytokines are
also involved in eosinophil migration induced by SAL or Sephadex, the
effect of pretreatment of saline-stimulated mast cell or macrophage
supernatants and Sephadex-stimulated mast cell supernatants with control
serum or antiserum against IL-1-beta, TNF-a, IL-5 or IL-8 was determined
(Fig. 5). Incubation of the
supernatants from saline- or
Sephadex-stimulated mast cells or macrophages with antibodies against
IL-1-beta and TNF-alpha had no effect on the subsequent eosinophil
migration. In contrast, incubation of supernatants from saline-stimulated
mast cells with antibodies against IL-5 or IL-8 abolished its ability to
induce eosinophil migration (Fig. 5a). The eosinophil chemotactic activity
of supernatants of saline-stimulated macrophages was only inhibited by the
antibody to IL-8 (Fig. 5b). In the supernatants from Sephadex-stimulated
mast cells only the antibody to IL-8 inhibited the eosinophil chemotactic
activity (Fig. 5c). All antisera were active at the concentration used
since they inhibited the ability of their respective cytokines to induce
eosinophil or neutrophil migration into the peritoneal cavity of naive rats
(data not shown). Until now, we have observed that, in addition to LTB4,
mast cells incubated with saline release IL-5 and IL-8, whereas macrophages
release IL-8. In the Sephadex model mast cells release LTB4 and IL-8.
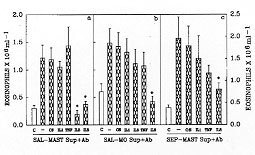
Figure 5: the
effects of antiserum against IL-1, TNF,
IL-5 and IL-8 on the eosinophil chemotactic activity of supernatants from
mast cells and macrophages incubated with saline or from mast cells
incubated with Sephadex. The bars represent the eosinophil migration
induced by the injection of the supernatants from mast cells (a) or
macrophages (b) previously incubated with saline. Panel c represents the
eosinophil migration induced by injection of the supernatant from mast
cells incubated with Sephadex. The supernatants were pretreated with PBS
(-), control serum (CS) or with IL-1, TNF, IL-5 or IL-8 antiserum before
injection. Eosinophil migration was evaluated 6 hr after injection of the
supernatants. The asterisks indicate significant differences between the
group incubated with PBS (-) and the groups treated with a given antiserum
(p<0.05; ANOVA followed by Bonfferoni's t test).
The next question was related to the mechanism which regulated eosinophil
migration induced by IL-5 or IL-8. As such, the administration of IL-5
induced a specific and dose-dependent eosinophil migration (3-25
ng/animal), which was already significant 6 hr after cytokine injection,
and remained high for up to 24 hr. The dose-response curve induced by IL-8
was bell-shaped. At the dose of 20 ng/rat, significant eosinophil migration
was observed, while at doses of 5, 10 and 40 ng/rat the eosinophil
migration observed did not differ from that induced by PBS. The migration
induced by 20 ng IL-8 only peaked 24 hr after injection of the cytokine and
returned to control levels after 48 hr (data not shown). Eosinophil
migration induced by IL-5 or IL-8 may have been blocked by the
pretreatments of the animals with MK 886 or dexamethasone. The IL-5-induced
eosinophil migration was also blocked by BW A4C, another 5-lipoxygenase
inhibitor (Fig. 6). We also
investigated the role of resident
peritoneal cells in IL-5 or IL-8-induced eosinophil migration. Eosinophil
migration induced by IL-5 or IL-8 was inhibited by 85 and 80%,
respectively, following prior depletion of the all resident cells by lavage
of the peritoneal cavity (data not shown). These data suggest that the
eosinophil migration induced by IL-5 or IL-8 is also dependent on the
resident peritoneal cells.
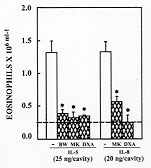
Figure 6: BW A4C, MK
886 and dexamethasone inhibited
the eosinophil migration induced by IL-5 or IL-8. The open bars show the
eosinophil migration induced by IL-5 (25 ng/cavity) or IL-8 (20 ng/cavity)
in PBS-pretreated animals. The hatched bars represent the eosinophil
migration in rats pretreated with MK 886 (MK, 1 mg/kg), BW A4C (BW, 20 mg/
kg) or dexamethasone (DXA, 0.5 mg/kg). Eosinophil migration was evaluated
24 hr after the injection of IL-5 or IL-8. The dashed line represents the
number of eosinophils in rats injected twice with PBS. The asterisks
indicate significant inhibition compared to the response in the nontreated
groups (-) (p<0.05; ANOVA followed by Bonfferoni's t test).
In conclusion, our results indicate that the eosinophil migration induced
by saline is dependent on resident mast cells and macrophages, whereas that
induced by Sephadex is only dependent on mast cells. Stimulated mast cells
release LTB4, IL-5 and IL-8 and macrophages release LTB4 and IL-8. IL-5 and
IL-8 released by the saline- or Sephadex-stimulated resident cells may act
in an autocrine fashion, thus potentiating the LTB4 release.
REFERENCES
Auriault C, Capron K, Cesarl IM, Capron A 1983. Enhancement of
eosinophil effector function by soluble factors released by S.
mansoni: role of protease. J Immunol 131: 464-470.
Barnes PJ, Chung KF, Page CP 1988. Inflammatory mediators in asthma.
Pharmacol Rev 40: 49-84.
Bascom R, Pipkorn U, Proud D, Dunnette S, Gleich GJ, Lichtenstein LM,
Naclerio RM 1989. Major basic protein and eosinophil-derived neurotoxin
concentrations in nasal-lavage fluid after antigen challenge: effect of
systemic corticosteroids and relationship to eosinophil influx. J
Allergy Clin Immunol 84: 338-346.
Berman JS, Weller PF 1992. Airway eosinophils and lymphocytes in
asthma. Am Rev Respir Dis 145: 1246-1248.
Capron M 1992. Dual function of eosinophils in pathogenesis and
protective immunity against parasites. Mem Inst Oswaldo Cruz 87
(Suppl V): 83-89.
Collins PD, Weg VB, Faccioli LH, Watson ML, Moqbel R, Williams TJ 1993.
Eosinophil accumulation induced by human interleukin-8 in the guinea pig
in vivo. Immunology 79: 312-318.
Cook NS 1988. The pharmacology of potassium channels and their
therapeutic potential. TIPS 91: 21-28.
Cook RM, Musgrove NRJ, Ashworth RF 1987. Activity of rat peritoneal
eosinophils following induction by different methods. Int Archs Allergy
Appl Immunol 83: 423-427.
Czarnetzki BM, Csato M 1989. Comparative studies of human eosinophil
towards platelet activating factor and leukotriene B4. Int Arch Allergy
Immunol 88: 191-193.
Davies WB, Fells GA, Sun X-H, Gadek JE, Venet A, Crystal RG 1984.
Eosinophil mediated injury to lung parenchyma cells and interstitial
matrix. A possible role for eosinophil in chronic inflammatory disorders
of the lower respiratory tract. J Clin Invest 74: 269-278.
Faccioli LH, Nourshargh S, Moqbel R, Williams FM, Sehmi R, Kay AB,
Williams TJ 1991. The accumulation of ^111In-eosinophils induced by
inflammatory mediators in vivo. Immunology 73: 222-227.
Gleich GJ, Flavahan NA, Fujisawa T, Vanhoutte PM 1988. The eosinophil
as a mediator of damage to respiratory epithelium: a model for bronchial
hyperreactivity. J Allergy Clin Immunol 81: 776-781.
Gleich GJ 1990. The eosinophil and bronchial asthma: current
understanding. J Allergy Clin Immunol 85: 422-435.
Hakansson L, Westerlund D, Venge P 1987. New method for the measurement
of eosinophil migration. J Leukocyte Biol 42: 689-696.
Holgate ST 1991. The mast cell and its function in allergic disease.
Clin Exp Allergy 21: 11-16.
Janiszewski J, Huizinga JD, Blennerhassett MG 1992. Mast cell ionic
channels: significance for stimulus - secretion coupling. Can J Physiol
Pharmacol 70: 1-7.
Jose PJ, Griffiths-Johnson DA, Collins PD, Wash DT, Moqbel R, Totty NF,
Truong O, Hsuan JJ, Willians TJ 1994. Eotaxin: A potent eosinophil
chemoattrac-tant cytokine detected in a guinea pig model of allergic
airways inflammation. J Exp Med 179: 881-887.
Kameyoshi Y, Dorschner A, Malet AI, Christophers E, Schroder J-M 1992.
Cytokine RANTES released by thrombin-stimulated platelets is a potent
attractant for human eosinophils. J Exp Med 176: 587-592.
Kay AB 1985. Eosinophils as effector cells in immunity and
hypersensitivity disorders. Clin Exp Immunol 62: 1-12.
Lee TH, Lane SJ 1992. The role of macrophages in the mechanisms of
airway inflammation in asthma. Am Rev Respir Dis 145:
S27-S30.
Leiferman KM, Ackerman SJ, Haugen HA, Venencie PY, Gleich GJ 1985.
Dermal deposition of eosinophil granule major basic protein in atopic
dermatitis. N Engl J Med 313: 282-285.
Lopez AF, Sanderson CJ, Gamble JR, Campbell HD, Young IG, Vadas MA
1988. Recombinant human interleukin 5 is selective activator of human
eosinophil function. J Exp Med 167: 219-224.
Maghni K, Blanchette F, Sirois P 1993. Induction of lung eosinophilia
and neutrophilia in guinea-pig following injection of sephadex beads.
Inflammation 17: 536-550.
Meacock SCR, Brandon DR, Smith W 1991. Interleukin-2 receptors on rat
eosinophils in adjuvant arthritis. Immunology 74: 169-171.
O'Donnell MC, Ackerman SJ, Gleich GJ, Thomas LL 1983. Activation of
mast cell and basophil histamine release by eosinophil granule major basic
protein. J Exp Med 157: 1981-1991.
Ogawa H, Kunkel SL, Fantone JC, Ward PA 1981. Comparative study of
eosinophil and neutrophil chemotaxis and enzyme release. Am J
Pathol 105: 149-155.
Raible DG, Schulman ES, Dimuzio J, Cardillo R, Post TJ 1992. Mast cell
mediators prostaglandin D2 and histamine activate human eosinophils. J
Immunol 148: 3536-3542.
Rand TH, Silberstein DS, Kornfeld H, Weller PF 1991. Human eosinophils
express functional interleukin 2 receptor. J Clin Invest 88:
825-832.
Resnick MB, Weller PF 1993 Mechanism of eosinophil recruitment. Am J
Resp Cell Mol Biol 8: 349-355.
Schriber RA, Zucker-Franklin D 1974. A method for the induction of
blood eosinophils with simple protein antigens. Cell Immunol
14: 470-474.
Sehmi R, Wardlaw AJ, Cromwell O, Kurihara K, Waltmann P, Kay AB 1992.
Interleukin 5 selectively enhances the chemotactic response of eosinophils
obtained from normal but not eosinophil subjects. Blood 79:
2952-2959.
Spicer BA, Baker RC, Halt PA, Laycock SM, Smith H 1990. The effecs of
drugs on Sephadex-induced eosinophilia and lung hyper-responsiveness.
Br J Pharmacol 101: 821-828.
Sun X-H, Davis WB, Fukuda Y, Ferrans VJ, Crystal RG 1985. Experimental
polymyxin B-induced interstitial lung disease characterised by an
accumulation of cytotoxic eosinophils in the alveolar structures. Am
Rev Resp Dis 131: 103-108.
Walls RS 1977. Eosinophil response to alum adjuvant: involvement of T
cells in non-antigen-dependent mechanisms. Proc Soc exp Biol Med
156: 431-435.
Copyright 1997 Fundacao Oswaldo Cruz - Fiocruz
The following images related to this document are available:
Photo images
[oc97188d.jpg]
[oc97188c.jpg]
[oc97188a.jpg]
[oc97188f.jpg]
[oc97188b.jpg]
[oc97188e.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}